Department of Bioengineering and Therapeutic Sciences, University of California, San Francisco, San Francisco, United States.
Biophysics Graduate Program, University of California San Francisco, San Francisco, United States.
Elife. 2022 Mar 21;11:e74114. doi: 10.7554/eLife.74114.
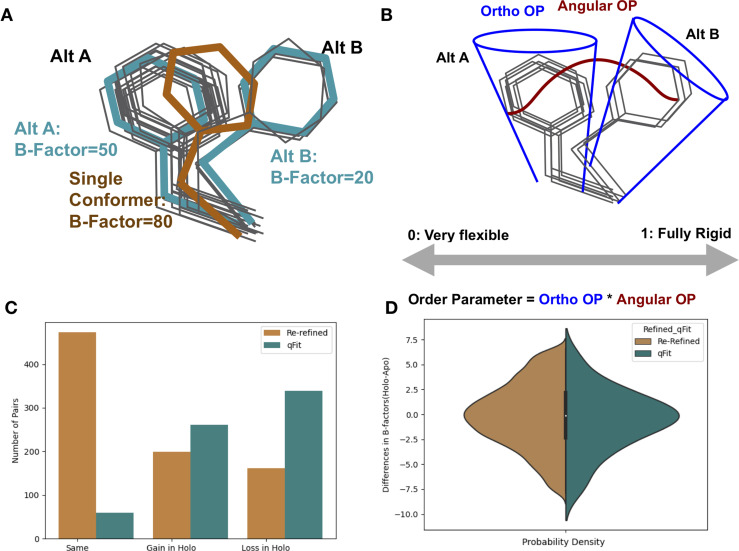
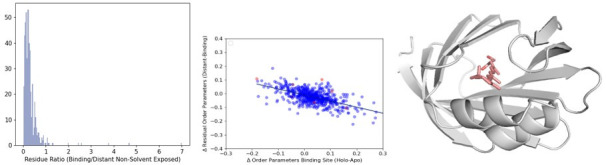
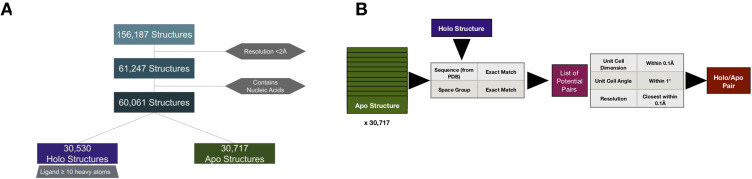
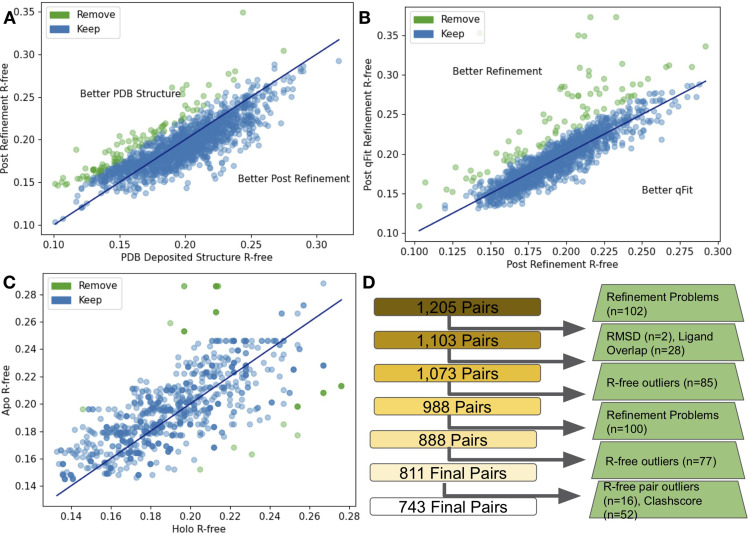
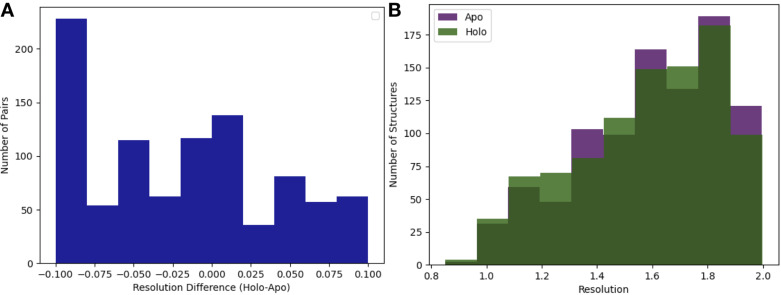
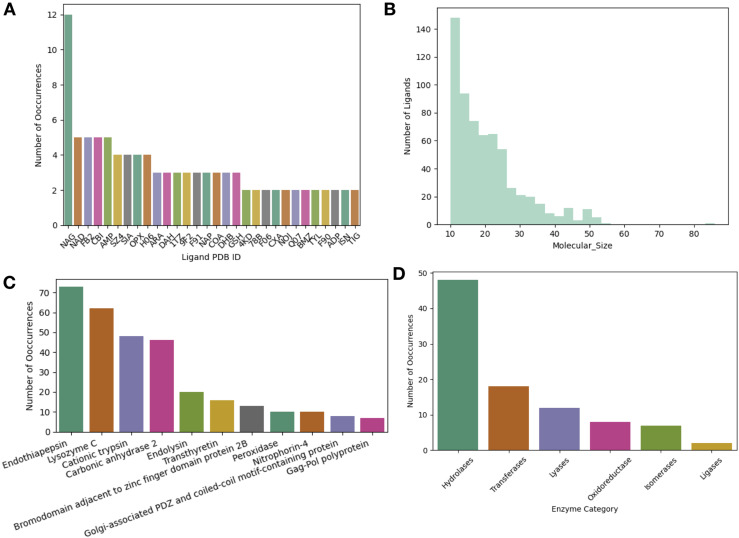
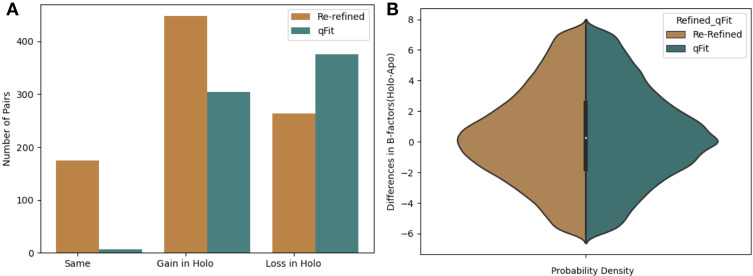
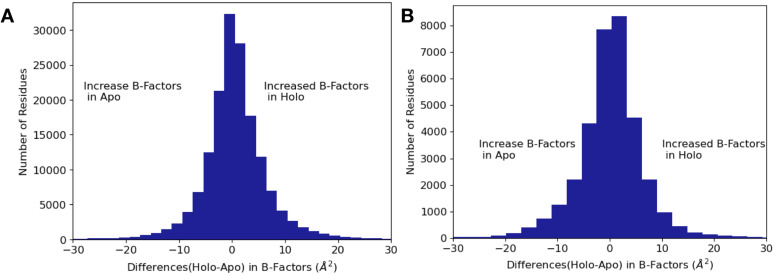
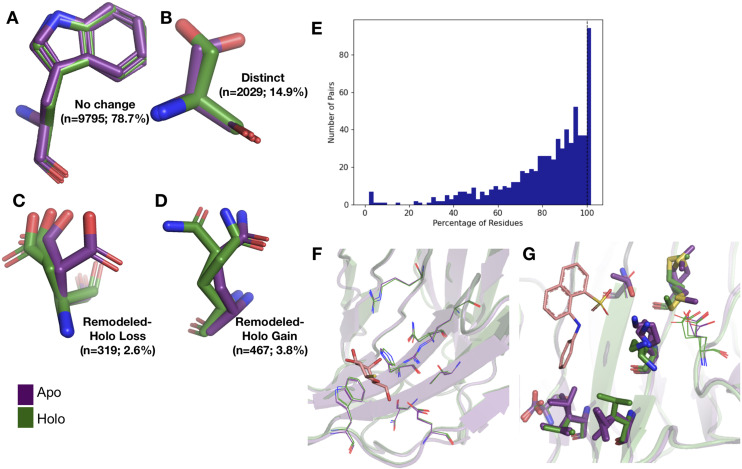
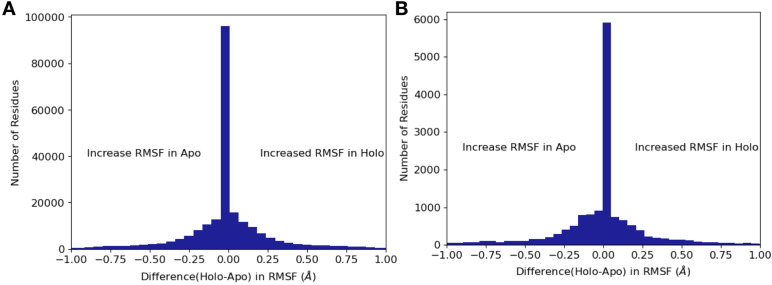
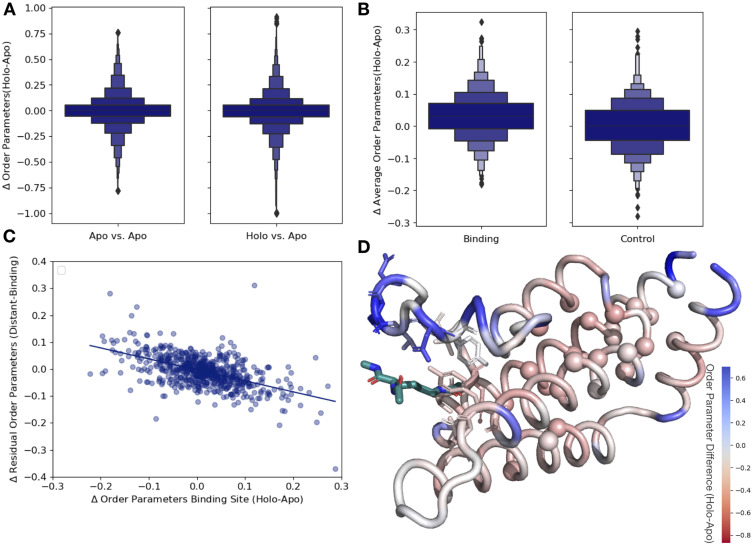
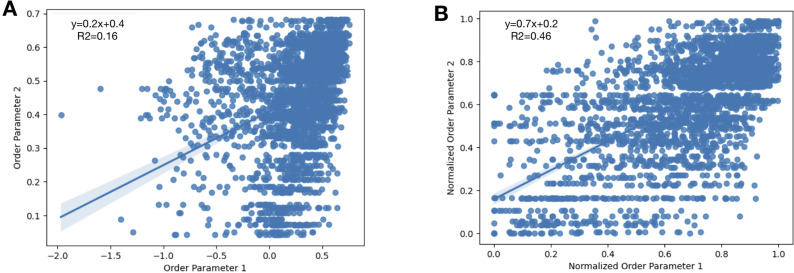
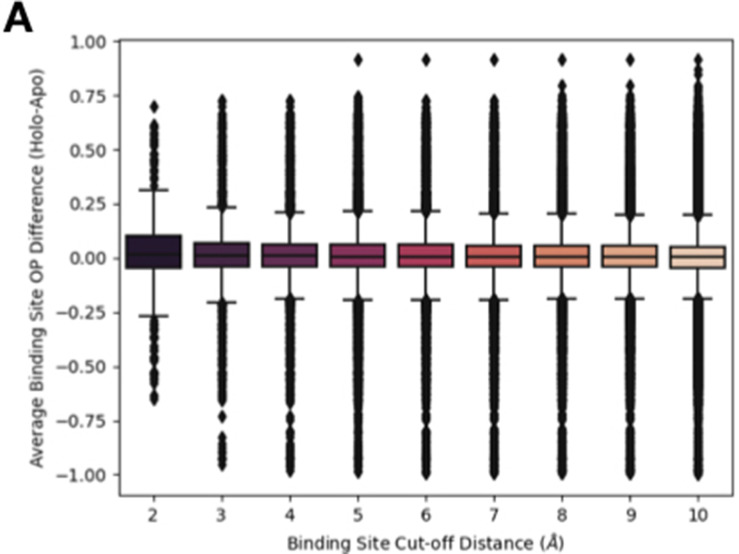
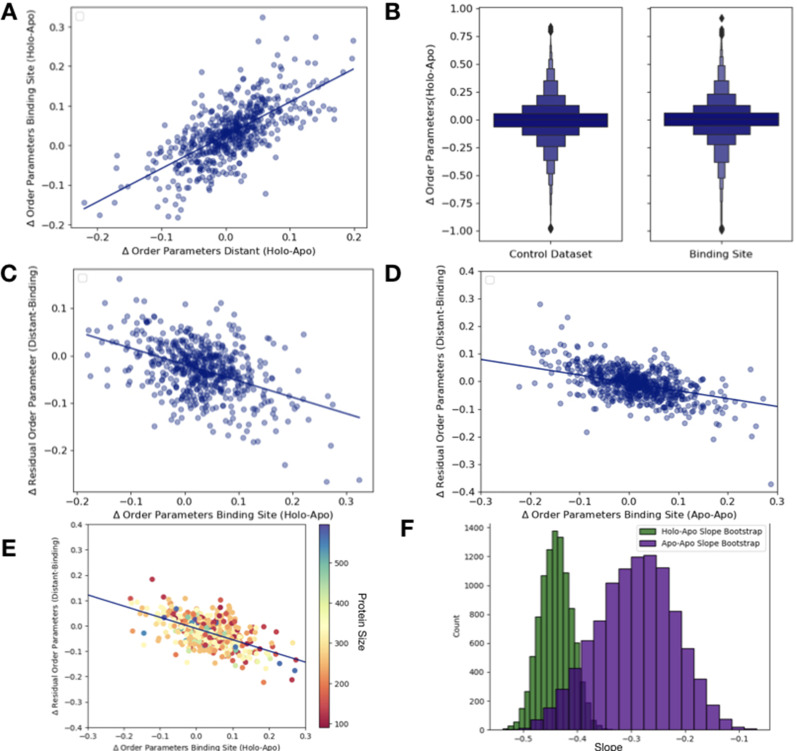
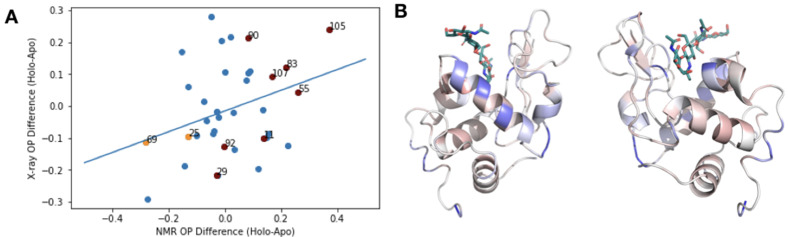
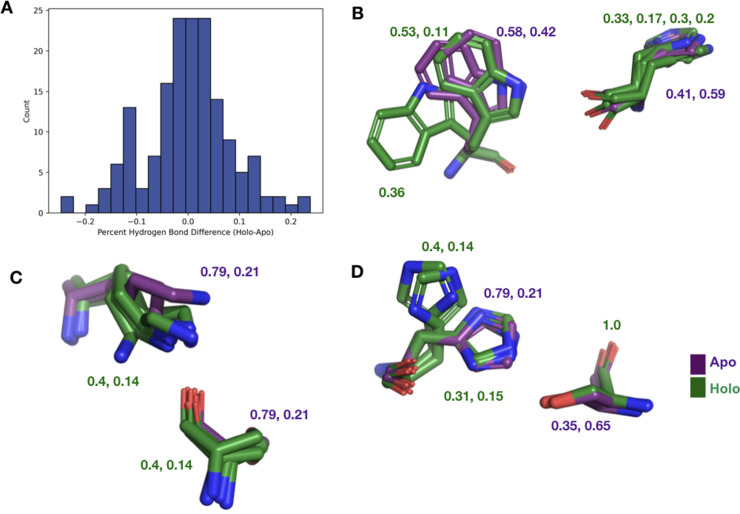
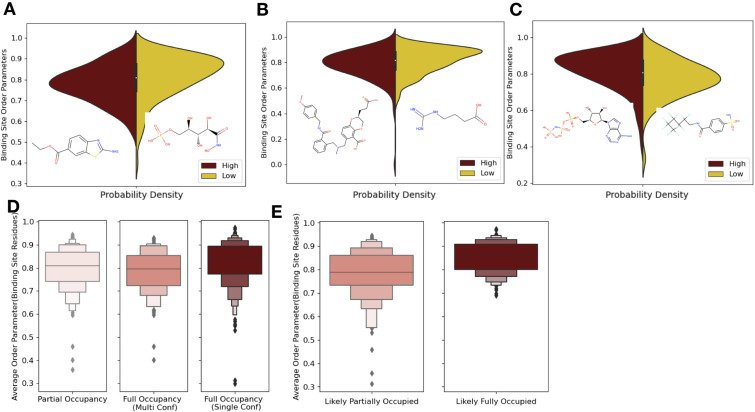
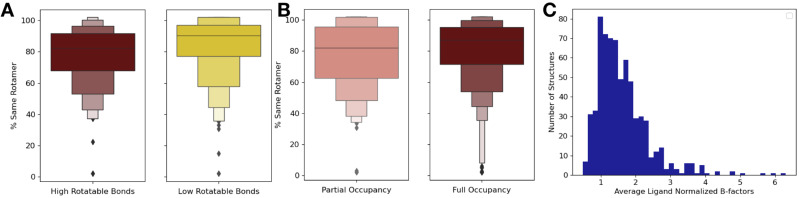
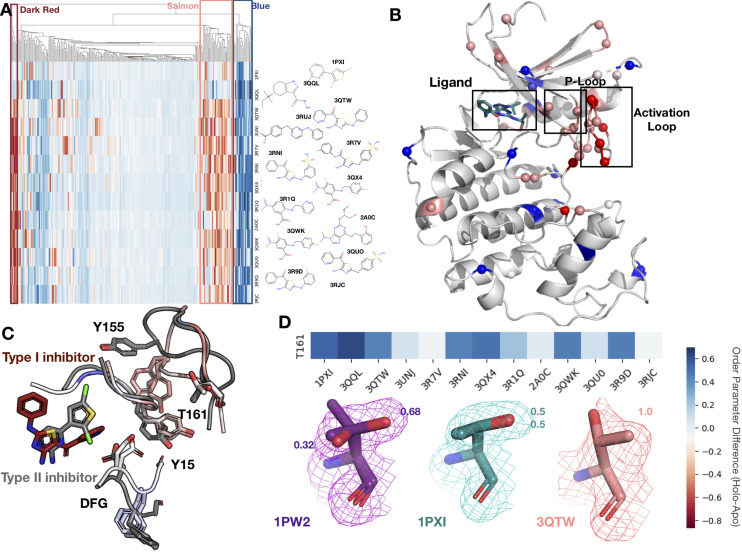
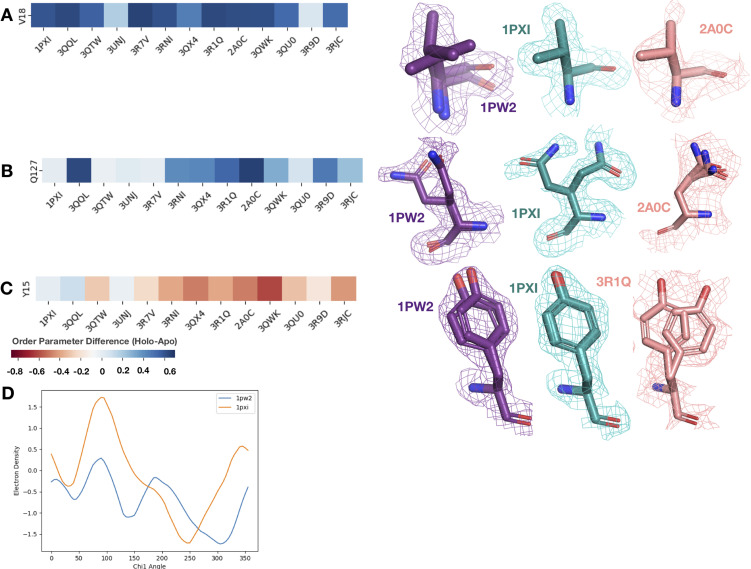
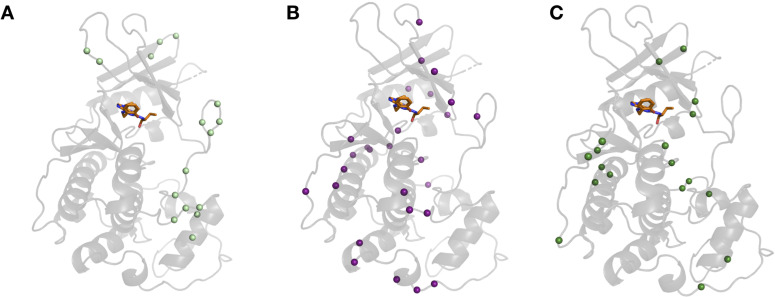
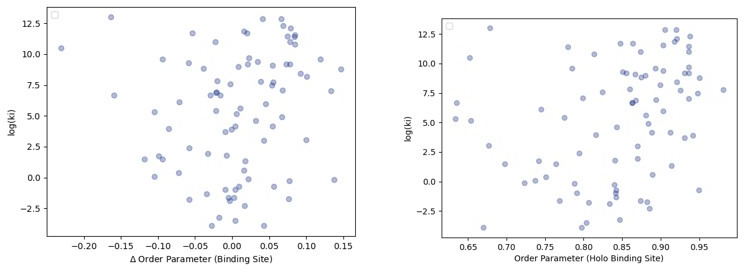
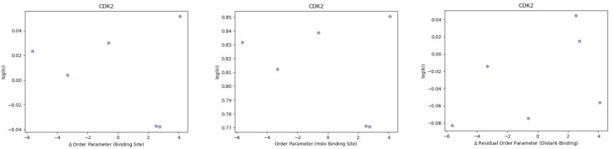
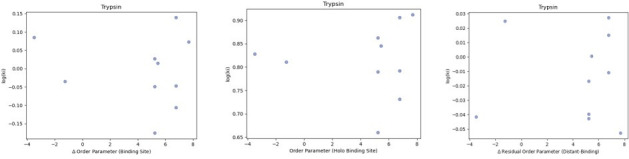
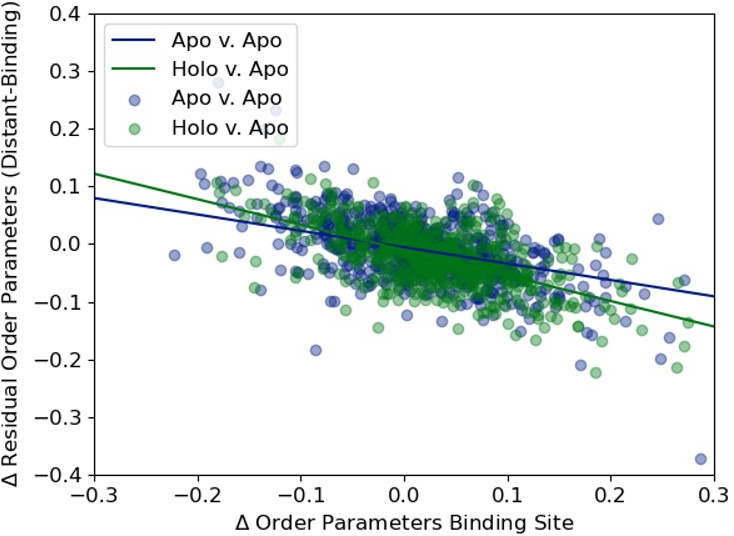
While protein conformational heterogeneity plays an important role in many aspects of biological function, including ligand binding, its impact has been difficult to quantify. Macromolecular X-ray diffraction is commonly interpreted with a static structure, but it can provide information on both the anharmonic and harmonic contributions to conformational heterogeneity. Here, through multiconformer modeling of time- and space-averaged electron density, we measure conformational heterogeneity of 743 stringently matched pairs of crystallographic datasets that reflect unbound/apo and ligand-bound/holo states. When comparing the conformational heterogeneity of side chains, we observe that when binding site residues become more rigid upon ligand binding, distant residues tend to become more flexible, especially in non-solvent-exposed regions. Among ligand properties, we observe increased protein flexibility as the number of hydrogen bonds decreases and relative hydrophobicity increases. Across a series of 13 inhibitor-bound structures of CDK2, we find that conformational heterogeneity is correlated with inhibitor features and identify how conformational changes propagate differences in conformational heterogeneity away from the binding site. Collectively, our findings agree with models emerging from nuclear magnetic resonance studies suggesting that residual side-chain entropy can modulate affinity and point to the need to integrate both static conformational changes and conformational heterogeneity in models of ligand binding.
虽然蛋白质构象异质性在许多方面都发挥着重要作用,包括配体结合,但它的影响一直难以量化。大分子 X 射线衍射通常采用静态结构进行解释,但它可以提供关于构象异质性的非谐和及调和贡献的信息。在这里,我们通过对时间和空间平均电子密度的多构象建模,测量了 743 对严格匹配的晶体数据集的构象异质性,这些数据集反映了无配体/apo 和配体结合/holo 状态。在比较侧链构象异质性时,我们观察到当结合位点残基在配体结合时变得更加刚性时,远距离残基往往变得更加灵活,特别是在非溶剂暴露区域。在一系列 13 个 CDK2 抑制剂结合结构中,我们发现构象异质性与抑制剂特征相关,并确定构象变化如何将构象异质性的差异从结合位点传播开来。总的来说,我们的发现与核磁共振研究中出现的模型一致,表明残侧链熵可以调节亲和力,并指出需要在配体结合模型中整合静态构象变化和构象异质性。