Milarski K L, Morimoto R I
Proc Natl Acad Sci U S A. 1986 Dec;83(24):9517-21. doi: 10.1073/pnas.83.24.9517.
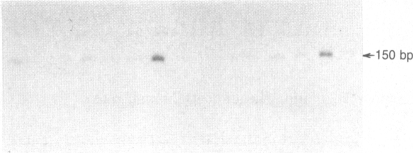
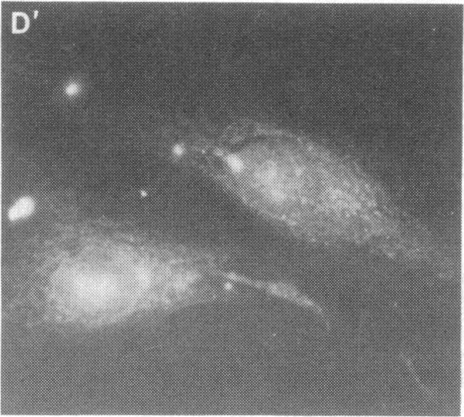
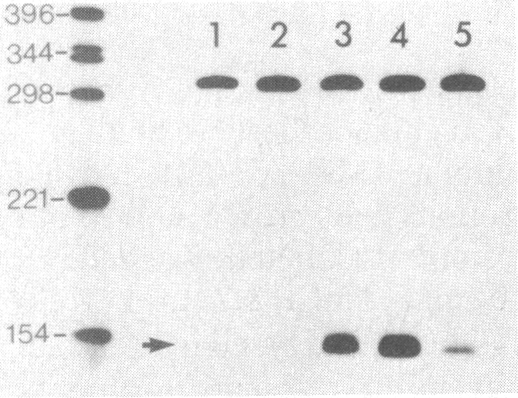
Expression of the major heat shock and stress-induced protein, HSP70, is under complex regulatory control in human cells. In addition to being induced by physiological stress such as heat shock or transition metals, the HSP70 gene is induced by serum stimulation and immortalizing products of the adenovirus E1A 13S and polyoma large tumor antigen genes. Here we show that expression of the human HSP70 gene is tightly regulated during the cell cycle. Using selective mitotic detachment, a noninductive method to obtain synchronous populations of HeLa cells, we show that levels of HSP70 mRNA rapidly increase 10- to 15-fold upon entry into S phase and decline by late S and G2. A transient increase in HSP70 synthesis is detected during early S phase. The subcellular localization of HSP70 varies throughout the cell cycle; the protein is diffusely distributed in the nucleus and cytoplasm in G1, localized in the nucleus in S, and again diffusely distributed in G2 cells. We suggest that the temporal pattern of HSP70 expression during S phase, the nuclear localization, and activation by trans-acting immortalizing proteins indicate a role for HSP70 in the nucleus of replicating cells.
主要热休克蛋白和应激诱导蛋白HSP70在人类细胞中受到复杂的调控。除了受热休克或过渡金属等生理应激诱导外,HSP70基因还可被血清刺激以及腺病毒E1A 13S和多瘤大肿瘤抗原基因的永生化产物诱导。在此我们表明,人类HSP70基因的表达在细胞周期中受到严格调控。利用选择性有丝分裂脱离这一非诱导性方法获得同步化的HeLa细胞群体,我们发现进入S期时HSP70 mRNA水平迅速增加10至15倍,并在S期末期和G2期下降。在S期早期检测到HSP70合成短暂增加。HSP70的亚细胞定位在整个细胞周期中有所不同;该蛋白在G1期在细胞核和细胞质中呈弥散分布,在S期定位于细胞核,在G2期细胞中又呈弥散分布。我们认为,HSP70在S期的表达时间模式、核定位以及被反式作用永生化蛋白激活表明其在复制细胞的细胞核中发挥作用。