Laboratory of Protein Metabolism, Graduate School of Pharmaceutical Sciences, The University of Tokyo, 7-3-1 Hongo, Bunkyo-ku, Tokyo 1130033, Japan.
Cells. 2022 May 7;11(9):1580. doi: 10.3390/cells11091580.
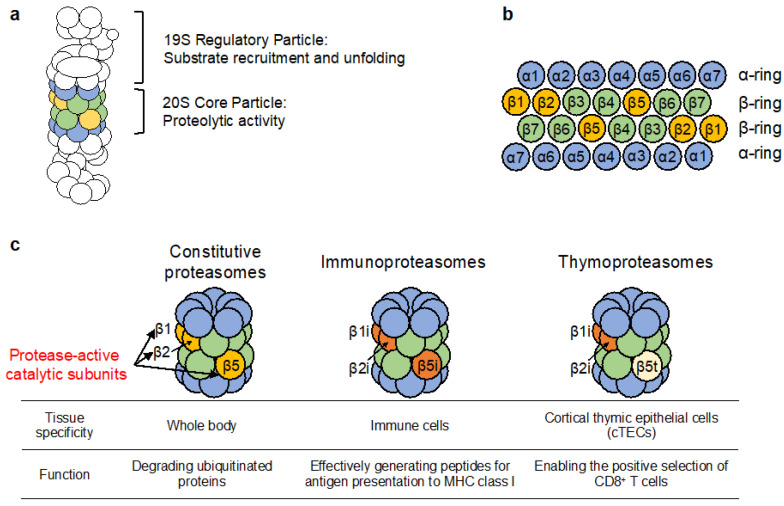
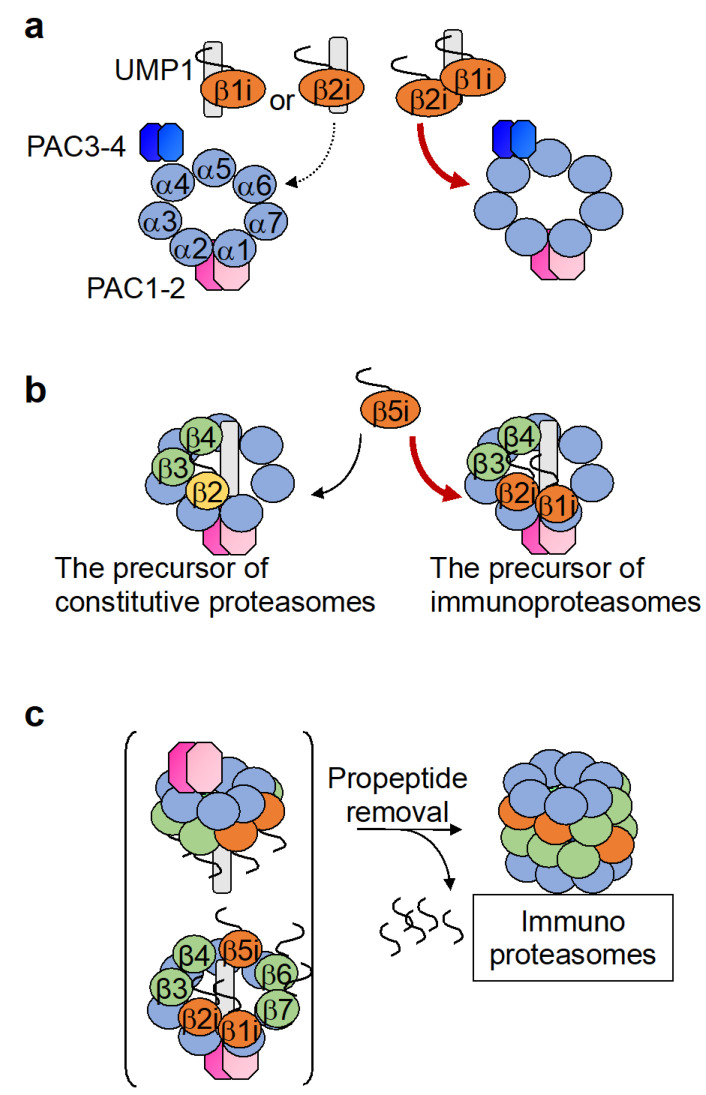
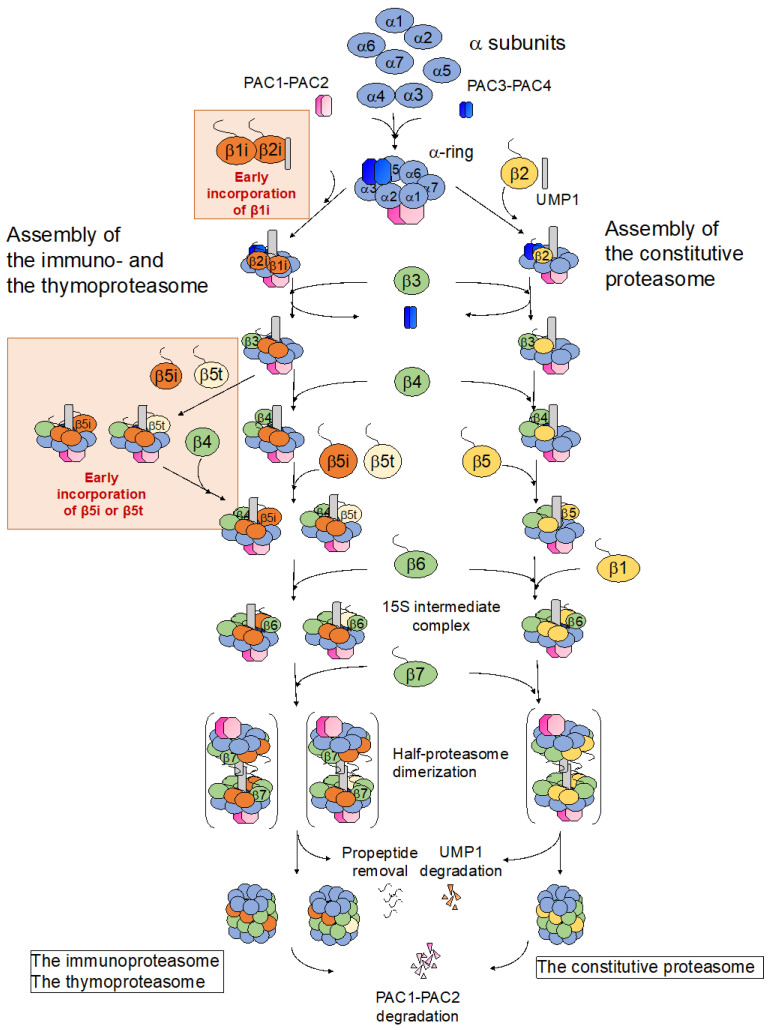
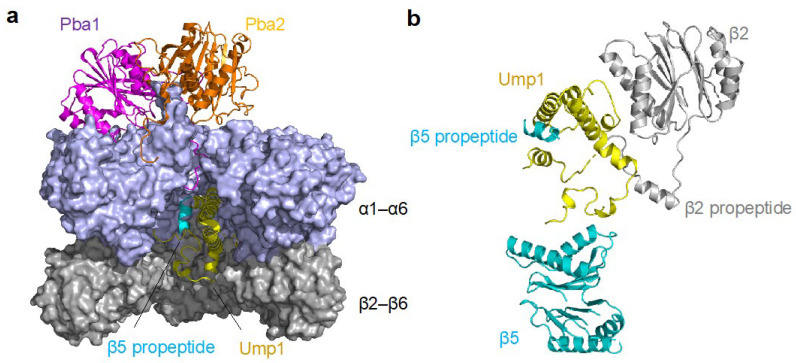
The proteasome is a large protein complex responsible for proteolysis in cells. Though the proteasome is widely conserved in all eukaryotes, vertebrates additionally possess tissue-specific proteasomes, termed immunoproteasomes and thymoproteasomes. These specialized proteasomes diverge from constitutive proteasomes in the makeup of their catalytic 20S core particle (CP), whereby the constitutive β1, β2, and β5 catalytic subunits are replaced by β1i, β2i, and β5i in immunoproteasomes, or β1i, β2i, and β5t in thymoproteasomes. However, as constitutive β1, β2, and β5 are also present in tissues and cells expressing immuno- and thymoproteasomes, the specialized proteasomes must be able to selectively incorporate their specific subunits. Here, we review the mechanisms governing the assembly of constitutive and specialized proteasomes elucidated thus far. Studies have revealed that β1i and β2i are added onto the α-ring of the CP prior to the other β subunits. Furthermore, β5i and β5t can be incorporated independent of β4, whereas constitutive β5 incorporation is dependent on β4. These mechanisms allow the immuno- and thymoproteasomes to integrate tissue-specific β-subunits without contamination from constitutive β1, β2, and β5. We end the review with a brief discussion on the diseases caused by mutations to the immunoproteasome and the proteins involved with its assembly.
蛋白酶体是一种负责细胞内蛋白水解的大型蛋白复合物。尽管蛋白酶体在所有真核生物中广泛保守,但脊椎动物还具有组织特异性的蛋白酶体,称为免疫蛋白酶体和胸腺蛋白酶体。这些特化的蛋白酶体在其催化 20S 核心颗粒 (CP) 的组成上与组成型蛋白酶体不同,其中组成型的β1、β2 和β5 催化亚基被免疫蛋白酶体中的β1i、β2i 和β5i 或胸腺蛋白酶体中的β1i、β2i 和β5t 取代。然而,由于组成型的β1、β2 和β5 也存在于表达免疫和胸腺蛋白酶体的组织和细胞中,因此特化的蛋白酶体必须能够选择性地掺入其特定的亚基。在这里,我们回顾了迄今为止阐明的调节组成型和特化蛋白酶体组装的机制。研究表明,β1i 和β2i 先于其他β 亚基添加到 CP 的α环上。此外,β5i 和β5t 可以独立于β4 掺入,而组成型β5 的掺入则依赖于β4。这些机制允许免疫和胸腺蛋白酶体在不被组成型β1、β2 和β5 污染的情况下整合组织特异性β-亚基。我们在简要讨论免疫蛋白酶体突变引起的疾病及其组装涉及的蛋白质后结束了综述。