Ludwig Institute for Cancer Research, Brussels, 1200, Belgium.
de Duve Institute, UCLouvain, Brussels, 1200, Belgium.
Sci Rep. 2020 Sep 25;10(1):15765. doi: 10.1038/s41598-020-71550-5.
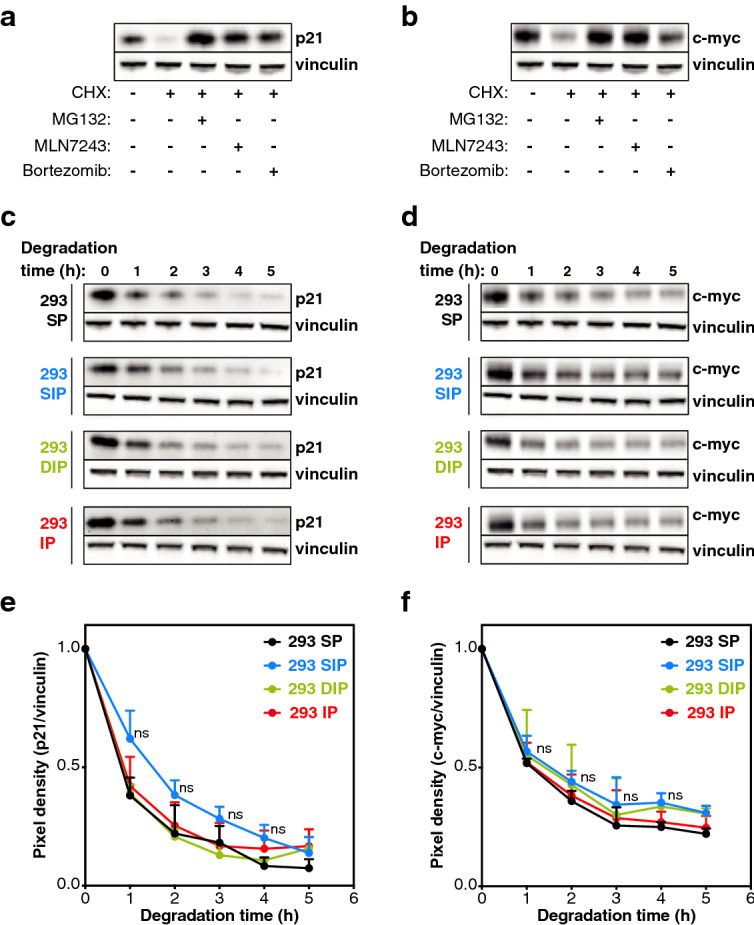
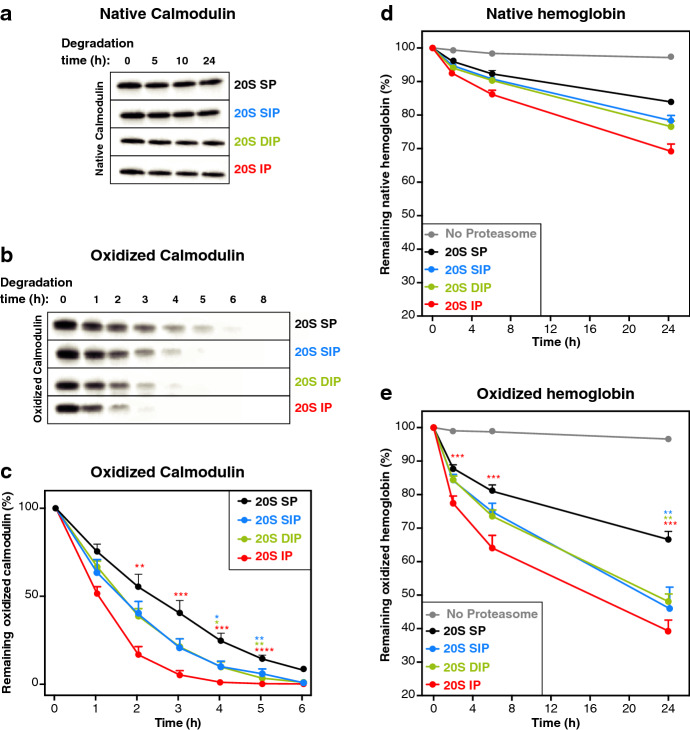
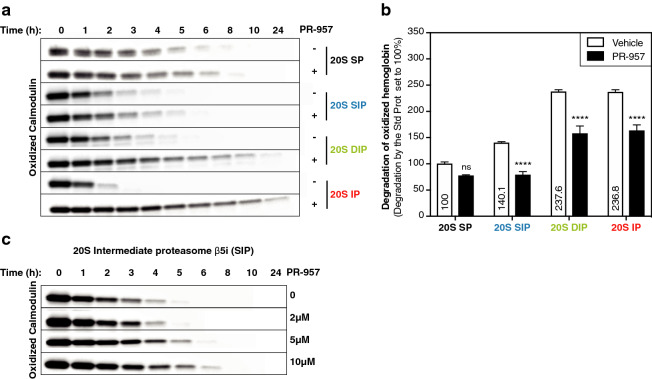
The proteasome is responsible for selective degradation of proteins. It exists in mammalian cells under four main subtypes, which differ by the combination of their catalytic subunits: the standard proteasome (β1-β2-β5), the immunoproteasome (β1i-β2i-β5i) and the two intermediate proteasomes (β1-β2-β5i and β1i-β2-β5i). The efficiency of the four proteasome subtypes to degrade ubiquitinated or oxidized proteins remains unclear. Using cells expressing exclusively one proteasome subtype, we observed that ubiquitinated p21 and c--myc were degraded at similar rates, indicating that the four 26S proteasomes degrade ubiquitinated proteins equally well. Under oxidative stress, we observed a partial dissociation of 26S into 20S proteasomes, which can degrade non-ubiquitinated oxidized proteins. Oxidized calmodulin and hemoglobin were best degraded in vitro by the three β5i-containing 20S proteasomes, while their native forms were not degraded. Circular dichroism analyses indicated that ubiquitin-independent recognition of oxidized proteins by 20S proteasomes was triggered by the disruption of their structure. Accordingly, β5i-containing 20S proteasomes degraded unoxidized naturally disordered protein tau, while 26S proteasomes did not. Our results suggest that the three β5i-containing 20S proteasomes, namely the immunoproteasome and the two intermediate proteasomes, might help cells to eliminate proteins containing disordered domains, including those induced by oxidative stress.
蛋白酶体负责选择性降解蛋白质。在哺乳动物细胞中,它存在于四种主要亚型中,这些亚型的区别在于其催化亚基的组合:标准蛋白酶体(β1-β2-β5)、免疫蛋白酶体(β1i-β2i-β5i)和两种中间蛋白酶体(β1-β2-β5i 和 β1i-β2-β5i)。四种蛋白酶体亚型降解泛素化或氧化蛋白的效率尚不清楚。使用仅表达一种蛋白酶体亚型的细胞,我们观察到泛素化的 p21 和 c--myc 以相似的速率降解,表明这四种 26S 蛋白酶体以相同的效率降解泛素化蛋白。在氧化应激下,我们观察到 26S 部分解离为 20S 蛋白酶体,后者可以降解非泛素化的氧化蛋白。体外,氧化钙调蛋白和血红蛋白被三种含有β5i 的 20S 蛋白酶体降解得最好,而其天然形式则不被降解。圆二色性分析表明,20S 蛋白酶体对氧化蛋白的泛素非依赖性识别是由其结构的破坏引发的。因此,含有β5i 的 20S 蛋白酶体降解未氧化的天然无序蛋白 tau,而 26S 蛋白酶体则不能。我们的结果表明,三种含有β5i 的 20S 蛋白酶体,即免疫蛋白酶体和两种中间蛋白酶体,可能有助于细胞消除含有无序结构域的蛋白质,包括由氧化应激诱导的蛋白质。