Department of Microbial Pathogenesis and Immunology, College of Medicine, Texas A&M Health Science Center, Bryan, United States.
Department of Electrical and Computer Engineering, Texas A&M University, College Station, United States.
Elife. 2022 May 19;11:e73625. doi: 10.7554/eLife.73625.
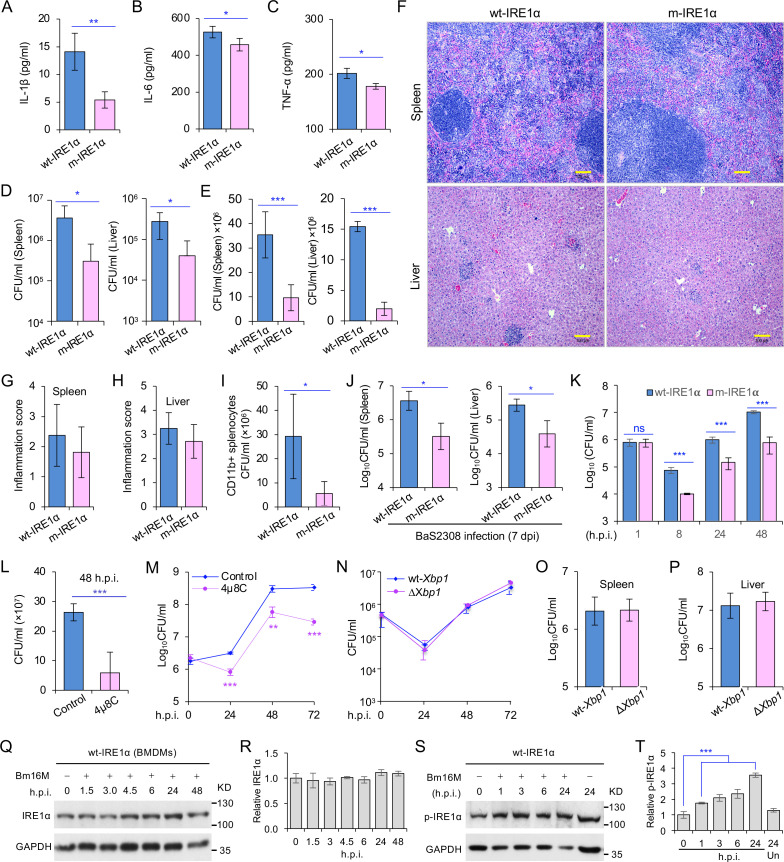
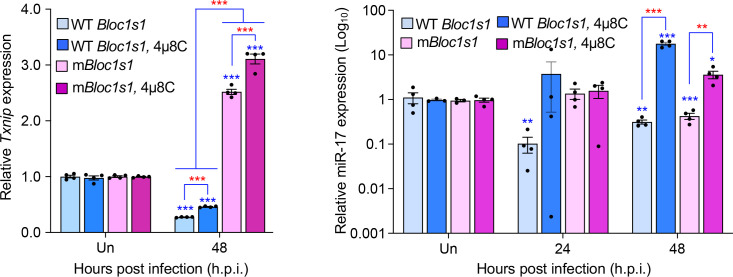
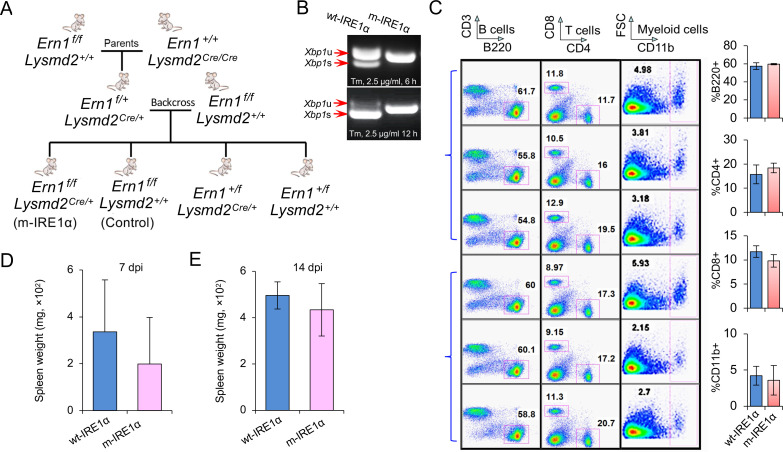
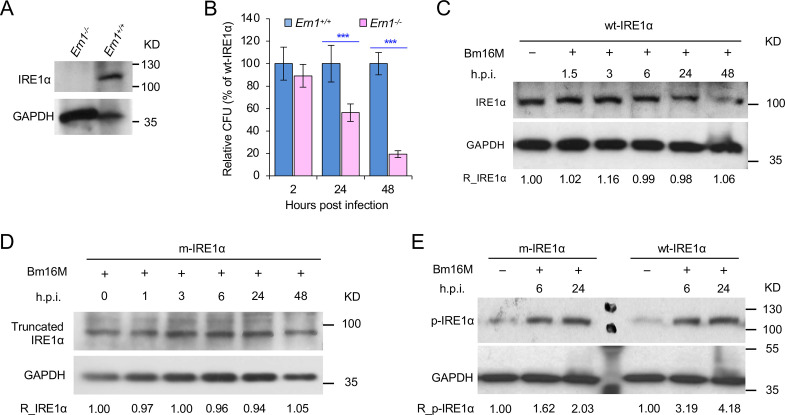
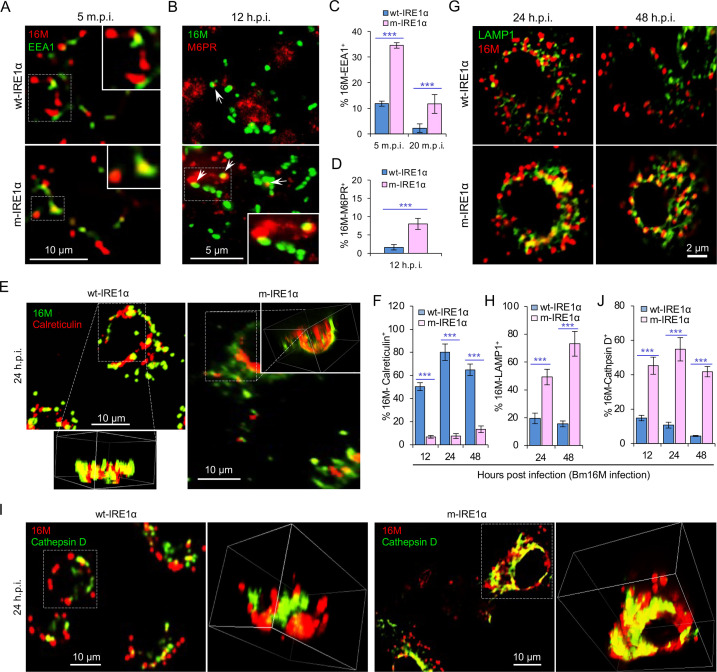
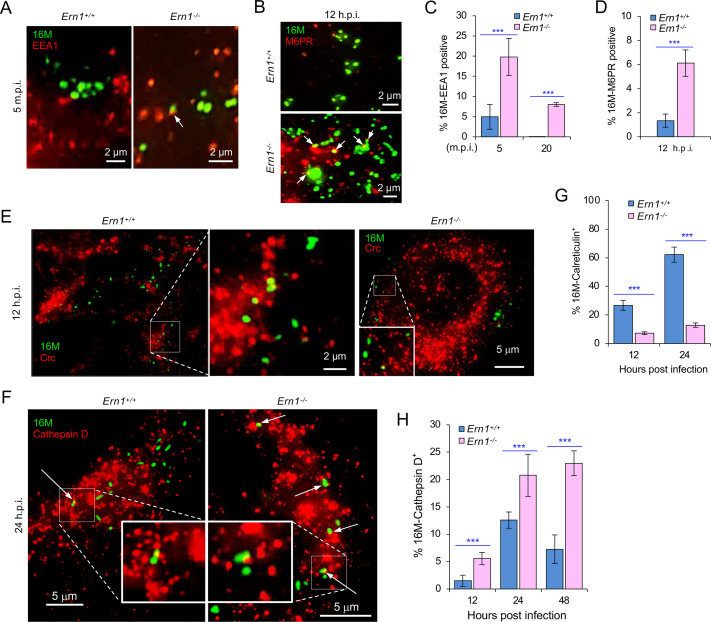
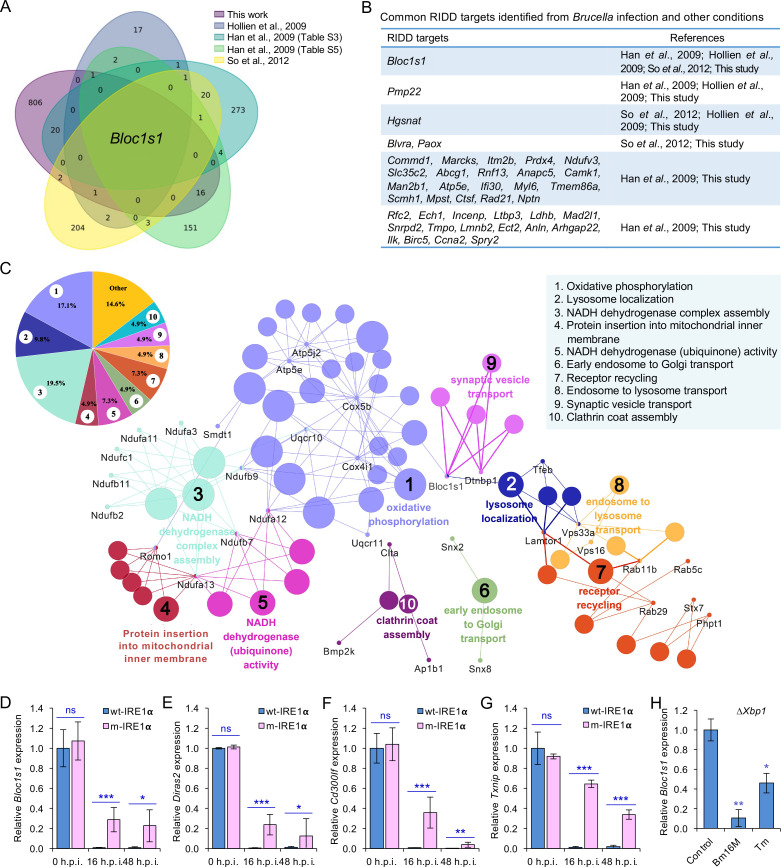
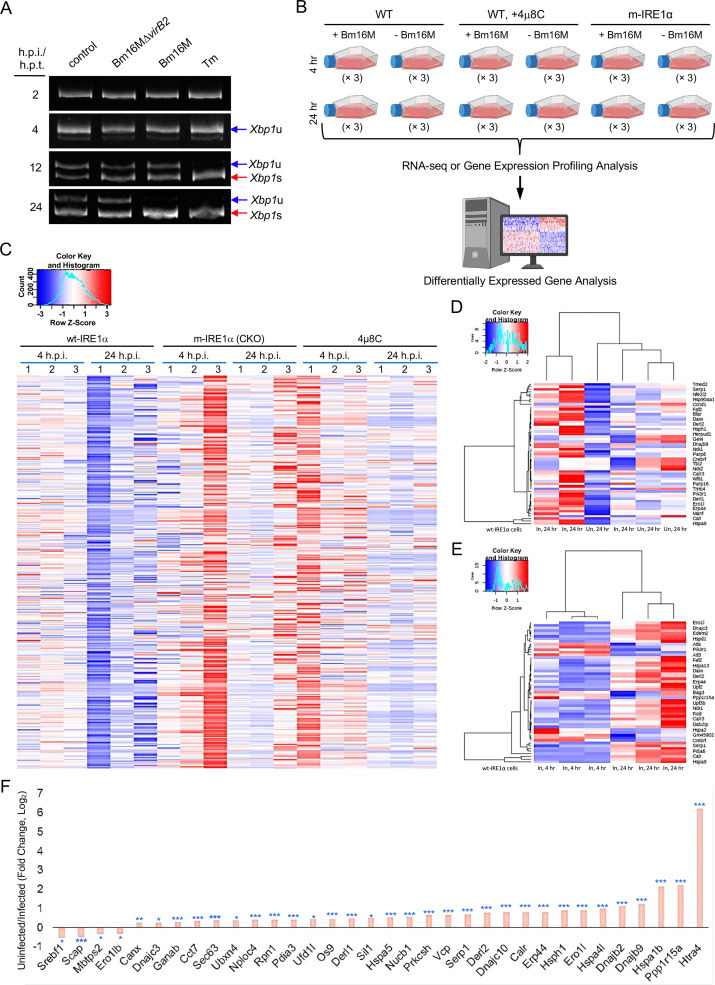
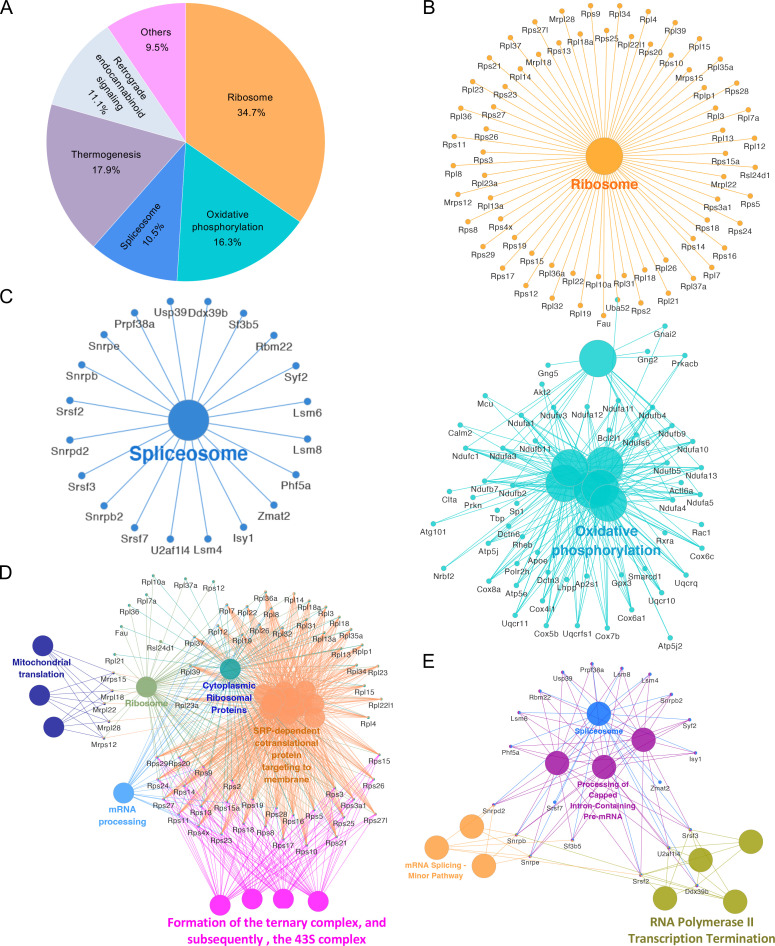
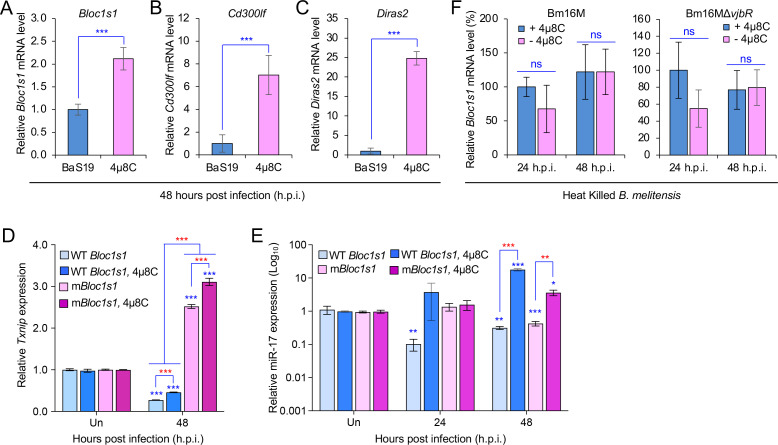
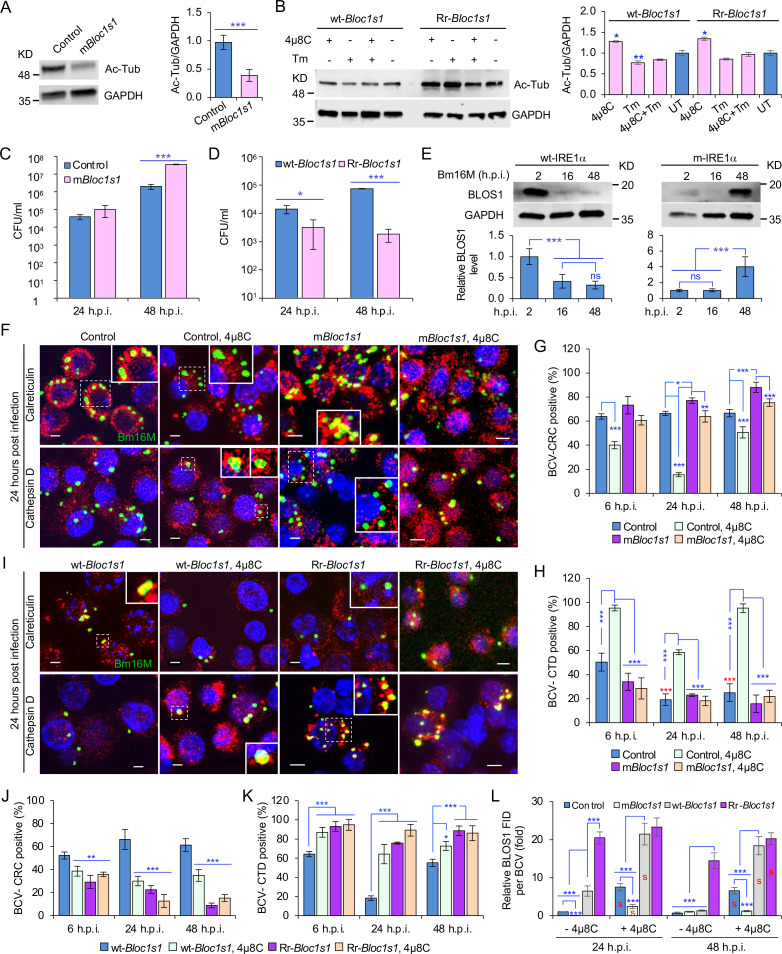
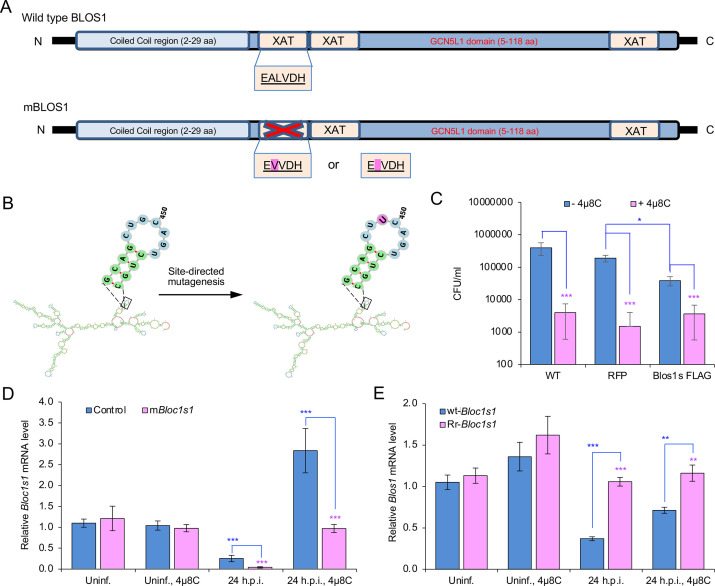
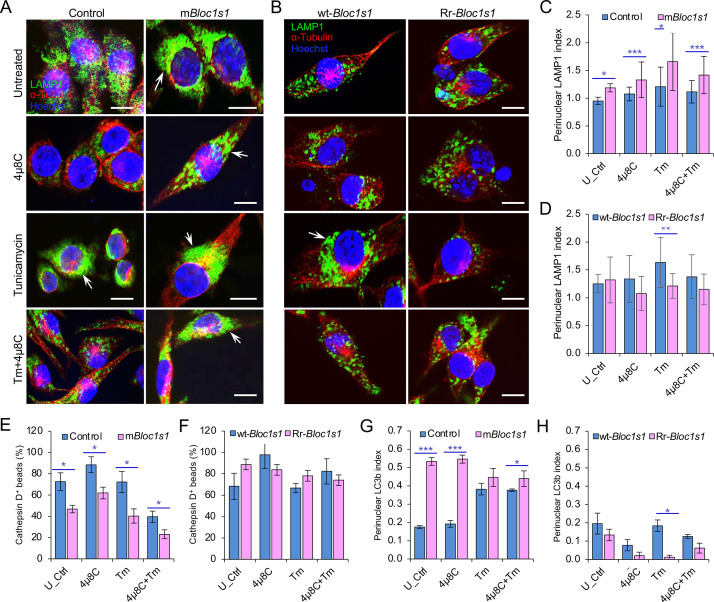
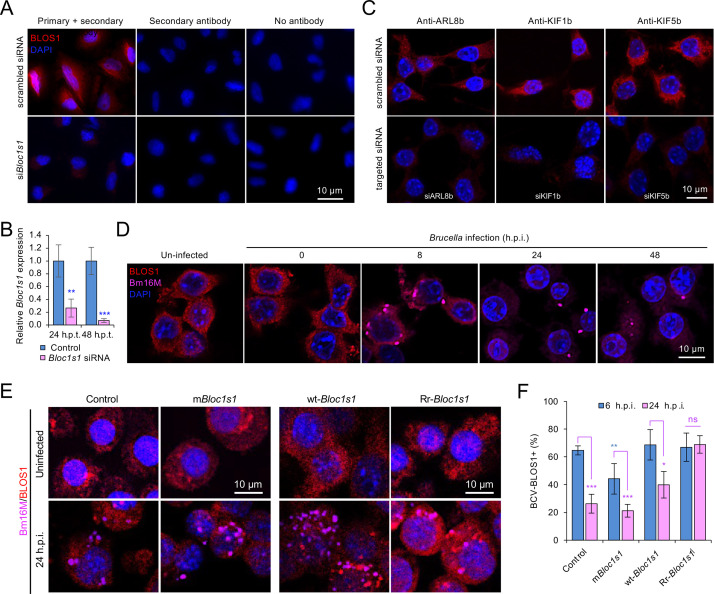
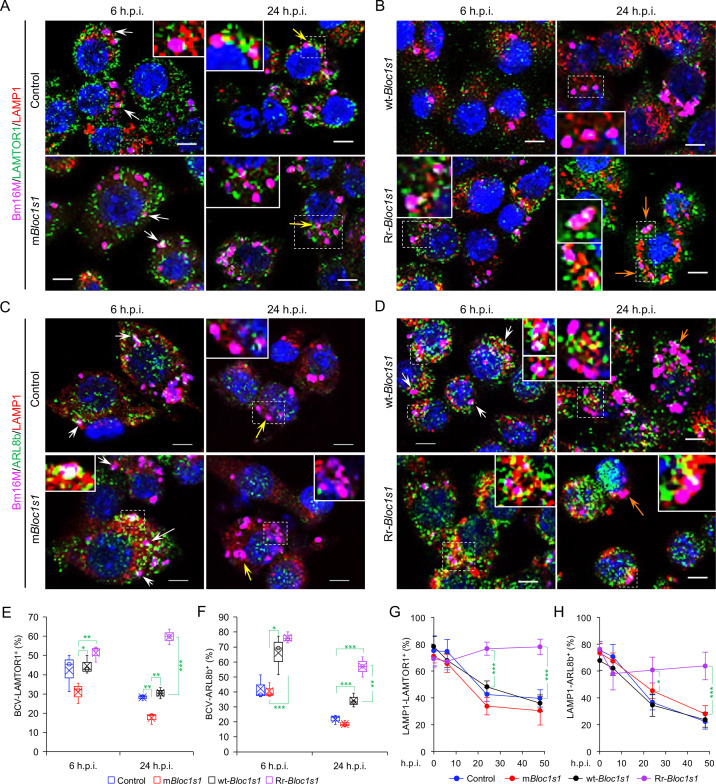
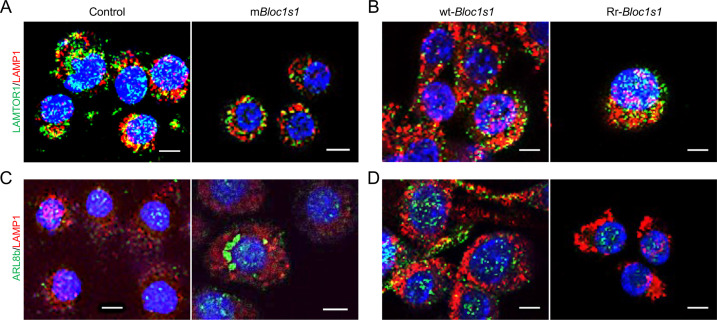
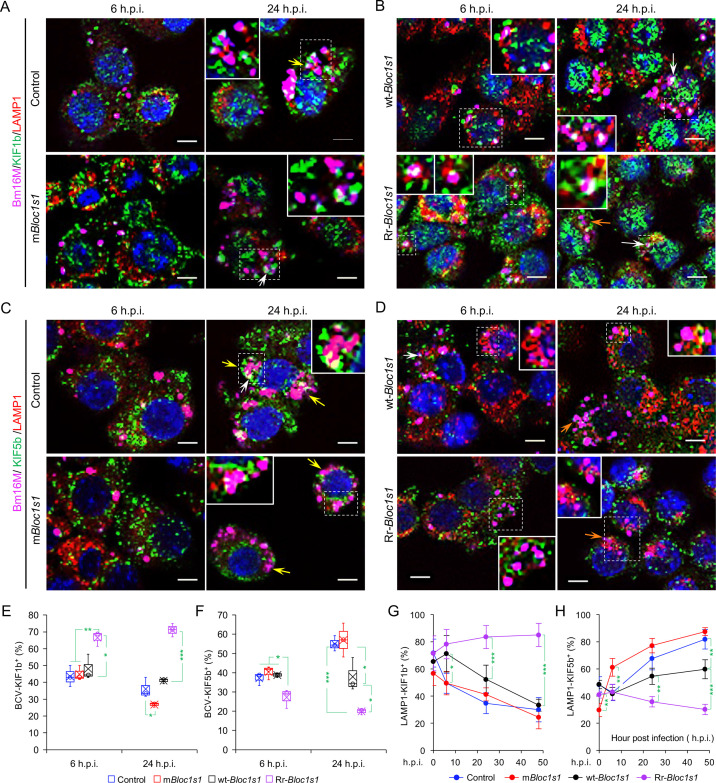
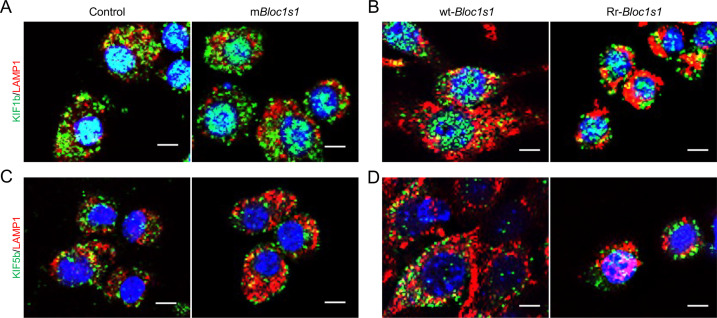
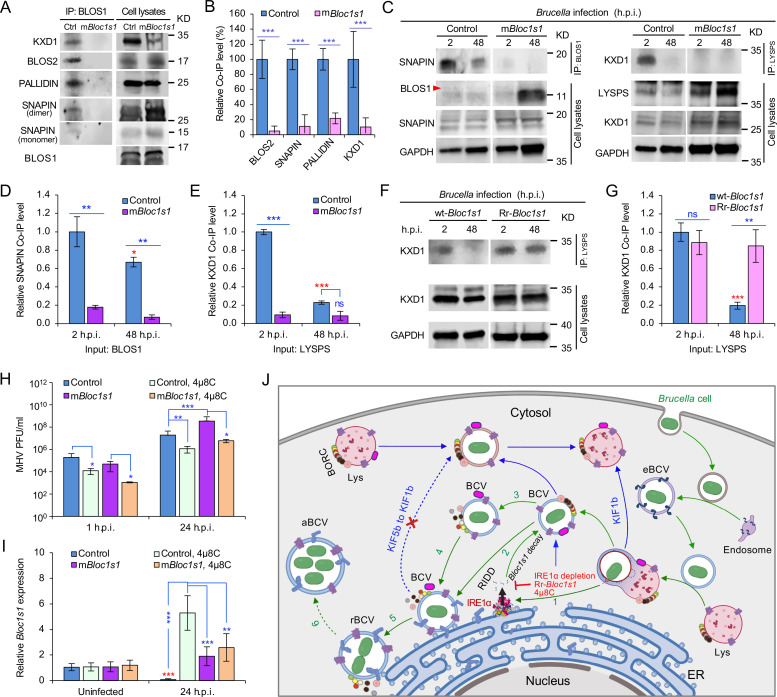
The phagocytosis and destruction of pathogens in lysosomes constitute central elements of innate immune defense. Here, we show that , the causative agent of brucellosis, the most prevalent bacterial zoonosis globally, subverts this immune defense pathway by activating regulated IRE1α-dependent decay (RIDD) of mRNA encoding BLOS1, a protein that promotes endosome-lysosome fusion. RIDD-deficient cells and mice harboring a RIDD-incompetent variant of IRE1α were resistant to infection. Inactivation of the gene impaired the ability to assemble BLOC-1-related complex (BORC), resulting in differential recruitment of BORC-related lysosome trafficking components, perinuclear trafficking of -containing vacuoles (BCVs), and enhanced susceptibility to infection. The RIDD-resistant variant maintains the integrity of BORC and a higher-level association of BORC-related components that promote centrifugal lysosome trafficking, resulting in enhanced BCV peripheral trafficking and lysosomal destruction, and resistance to infection. These findings demonstrate that host RIDD activity on BLOS1 regulates intracellular parasitism by disrupting BORC-directed lysosomal trafficking. Notably, coronavirus murine hepatitis virus also subverted the RIDD-BLOS1 axis to promote intracellular replication. Our work establishes BLOS1 as a novel immune defense factor whose activity is hijacked by diverse pathogens.
溶酶体吞噬和破坏病原体是先天免疫防御的核心要素。在这里,我们表明,布鲁氏菌病的病原体,也是全球最普遍的细菌性人畜共患病原,通过激活调节的 IRE1α 依赖的 mRNA 降解(RIDD)来颠覆这种免疫防御途径,该途径可降解编码 BLOS1 的 mRNA,BLOS1 是一种促进内体-溶酶体融合的蛋白。缺乏 RIDD 的细胞和携带 RIDD 失活的 IRE1α 变异体的小鼠对感染具有抗性。失活 基因会损害组装 BLOC-1 相关复合物(BORC)的能力,导致 BORC 相关溶酶体运输成分的差异募集、含 - 的液泡(BCVs)的核周运输以及对感染的易感性增加。RIDD 抗性 变异体保持 BORC 的完整性和更高水平的 BORC 相关成分的关联,促进离心溶酶体运输,从而增强 BCV 周边运输和溶酶体破坏,并对感染产生抗性。这些发现表明,宿主 BLOS1 上的 RIDD 活性通过破坏 BORC 定向的溶酶体运输来调节细胞内寄生虫病。值得注意的是,冠状病毒鼠肝炎病毒也颠覆了 RIDD-BLOS1 轴以促进细胞内复制。我们的工作确立了 BLOS1 作为一种新的免疫防御因子,其活性被多种病原体劫持。