Department of Bioengineering, University of Pennsylvania, Philadelphia, PA, USA.
Epigenetics Institute, Perelman School of Medicine, University of Pennsylvania, Philadelphia, PA, USA.
Nature. 2022 Jun;606(7915):812-819. doi: 10.1038/s41586-022-04803-0. Epub 2022 Jun 8.
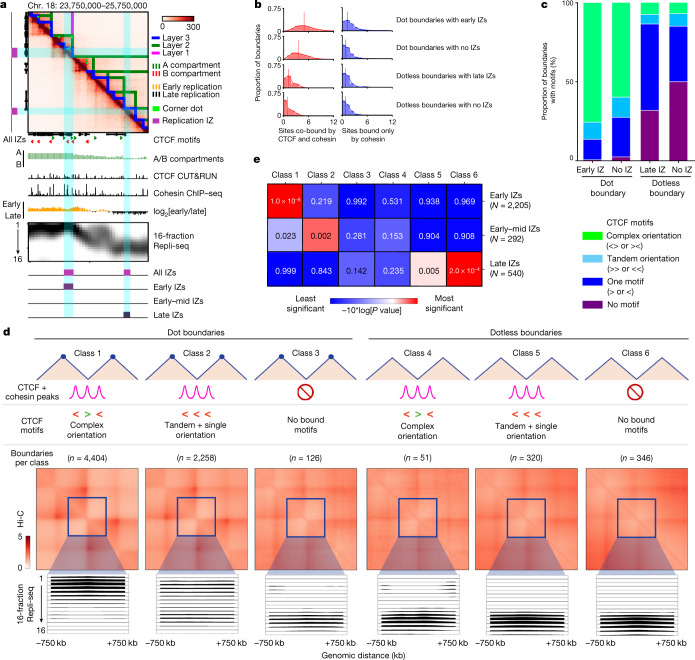
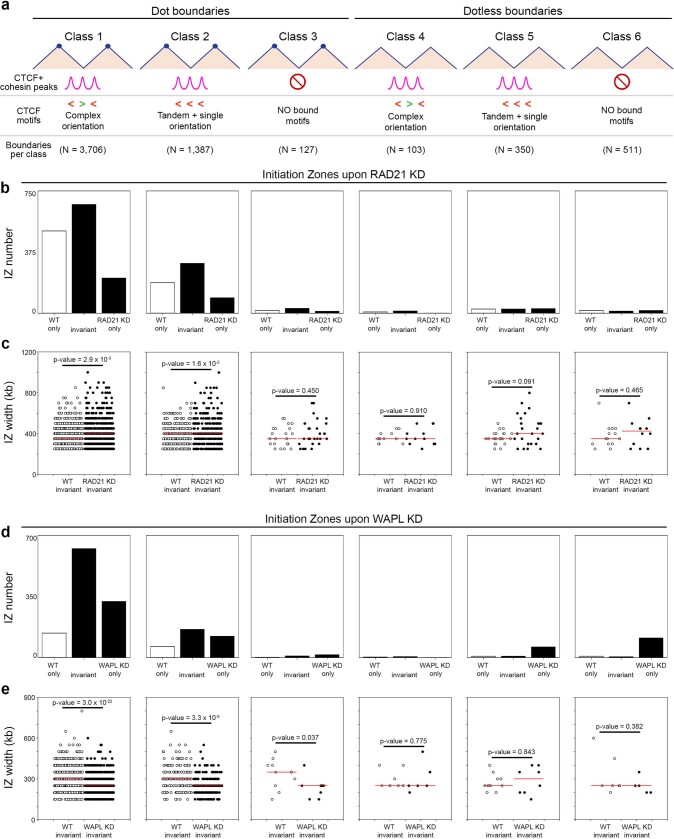
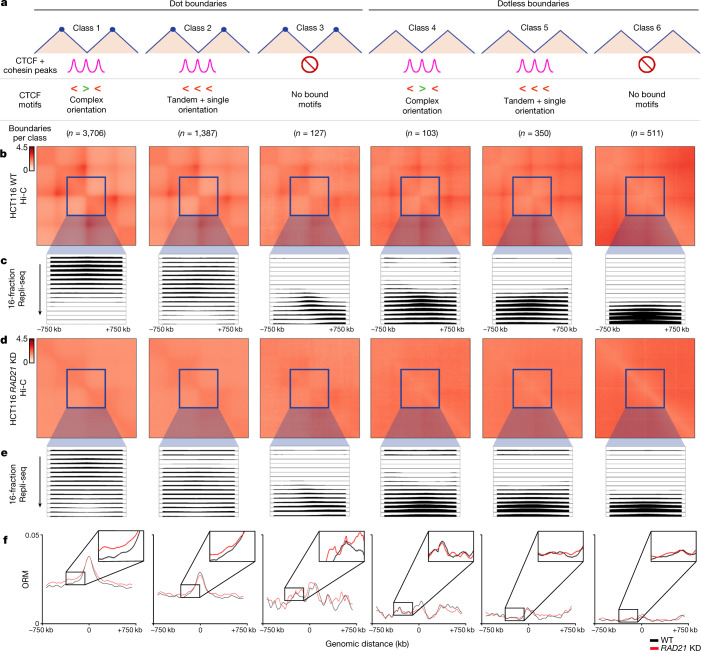
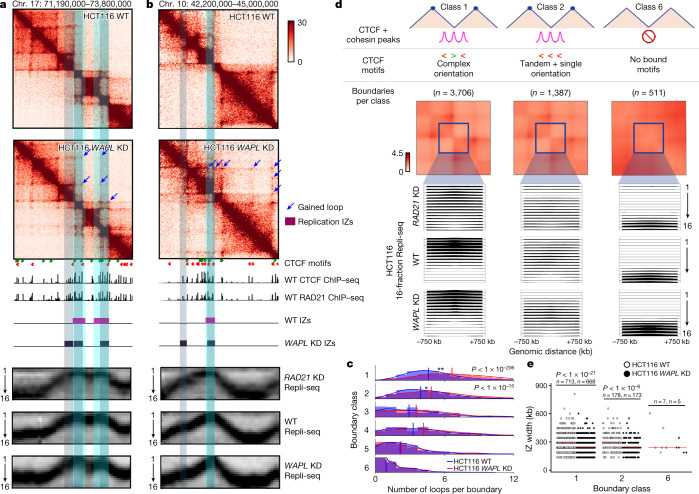
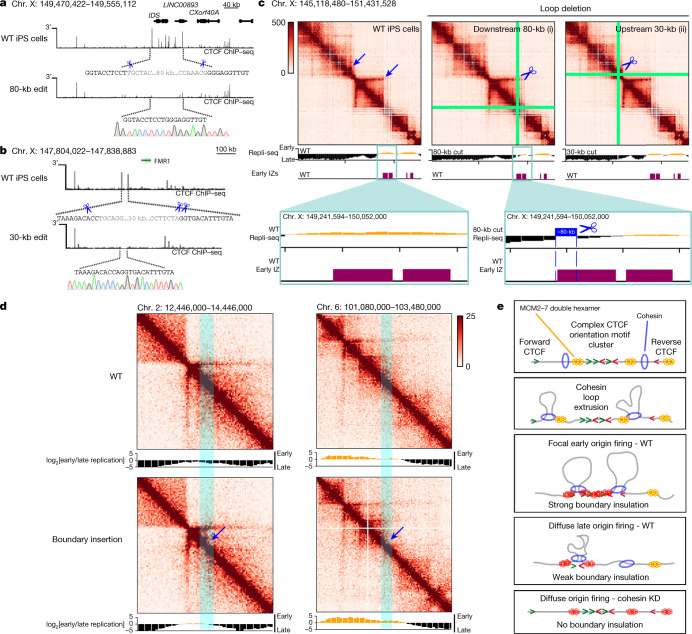
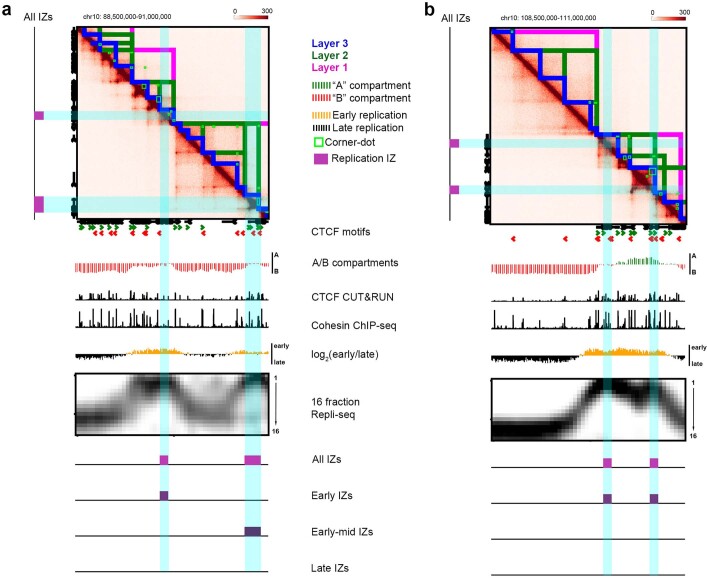
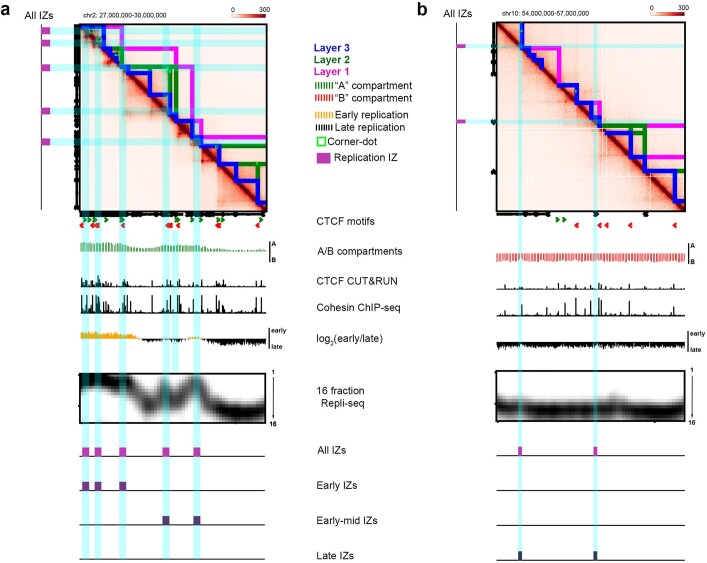
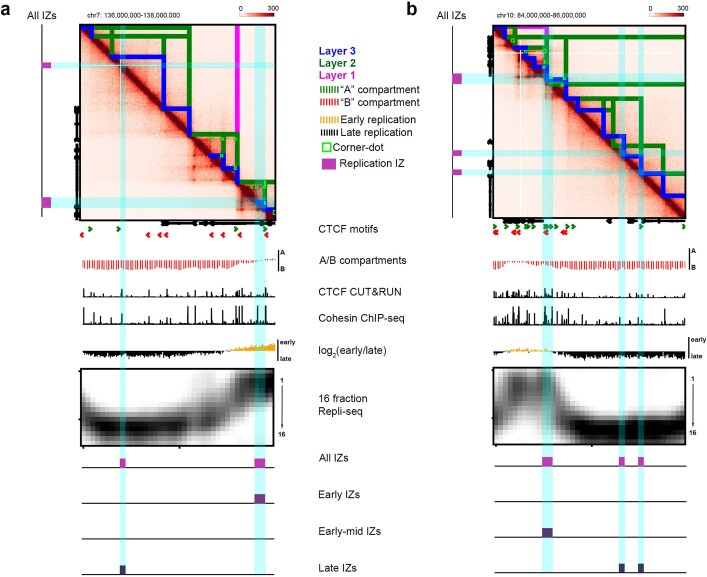
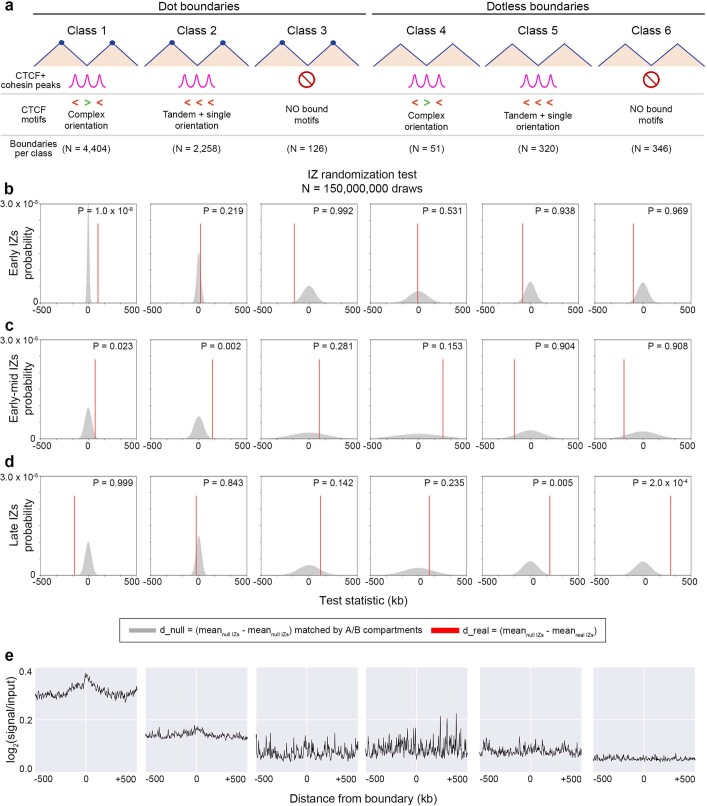
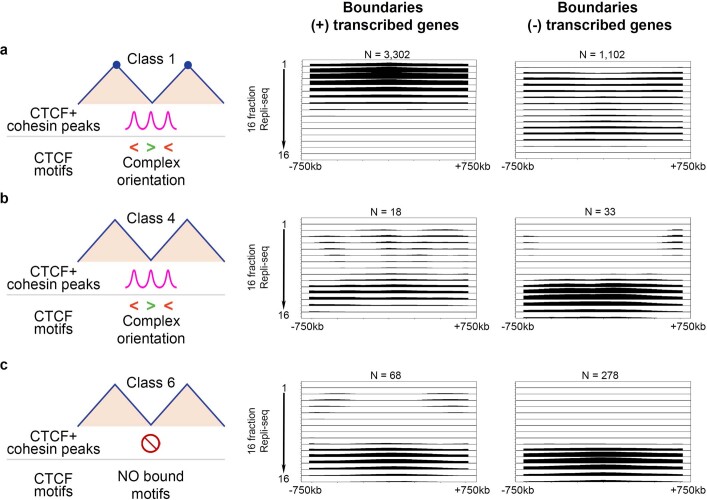
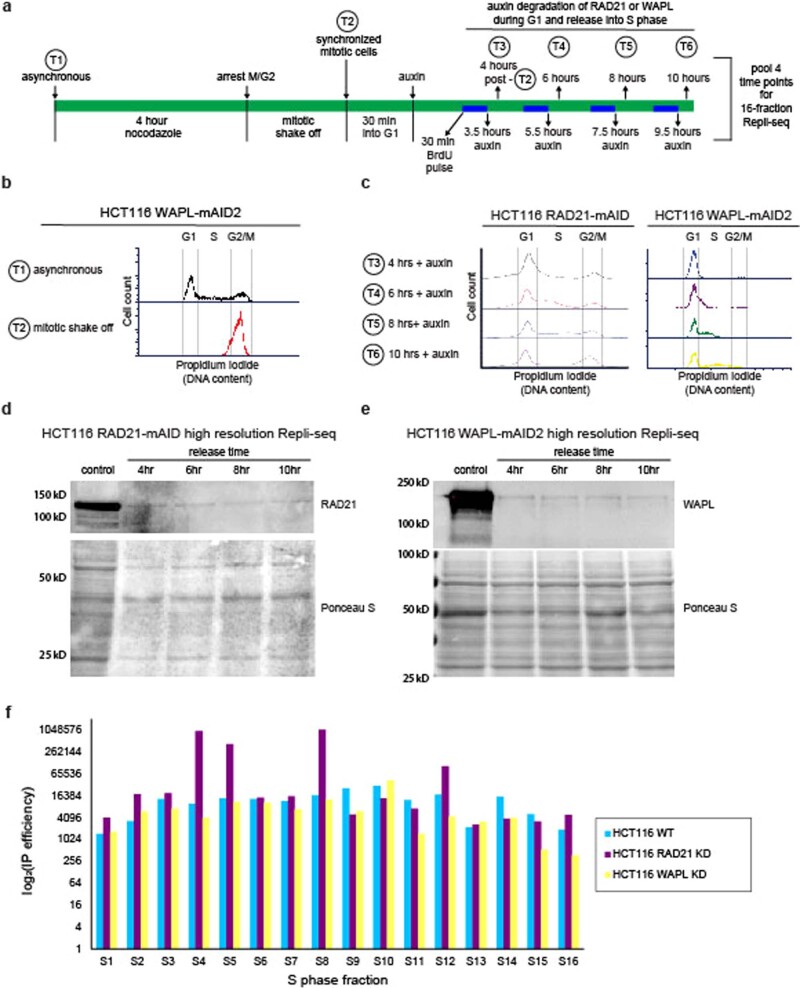
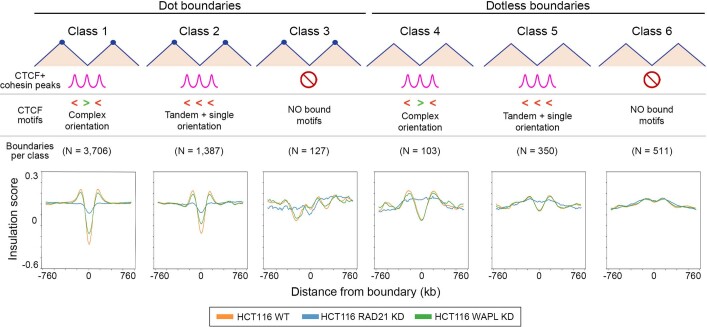
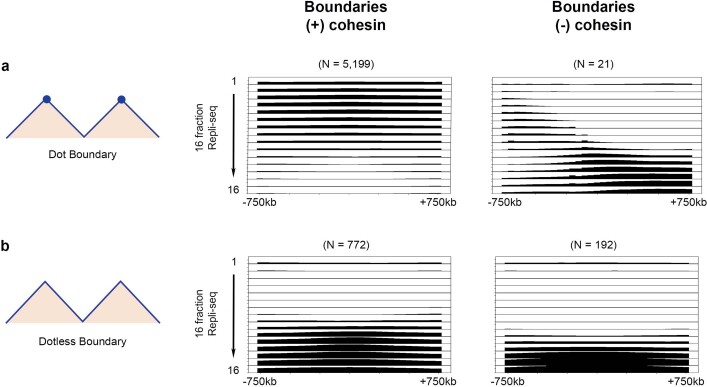
DNA replication occurs through an intricately regulated series of molecular events and is fundamental for genome stability. At present, it is unknown how the locations of replication origins are determined in the human genome. Here we dissect the role of topologically associating domains (TADs), subTADs and loops in the positioning of replication initiation zones (IZs). We stratify TADs and subTADs by the presence of corner-dots indicative of loops and the orientation of CTCF motifs. We find that high-efficiency, early replicating IZs localize to boundaries between adjacent corner-dot TADs anchored by high-density arrays of divergently and convergently oriented CTCF motifs. By contrast, low-efficiency IZs localize to weaker dotless boundaries. Following ablation of cohesin-mediated loop extrusion during G1, high-efficiency IZs become diffuse and delocalized at boundaries with complex CTCF motif orientations. Moreover, G1 knockdown of the cohesin unloading factor WAPL results in gained long-range loops and narrowed localization of IZs at the same boundaries. Finally, targeted deletion or insertion of specific boundaries causes local replication timing shifts consistent with IZ loss or gain, respectively. Our data support a model in which cohesin-mediated loop extrusion and stalling at a subset of genetically encoded TAD and subTAD boundaries is an essential determinant of the locations of replication origins in human S phase.
DNA 复制是通过一系列复杂的分子事件进行的,对于基因组的稳定性至关重要。目前,人类基因组中复制起始点的位置是如何确定的还不得而知。在这里,我们剖析了拓扑关联域(TAD)、亚 TAD 和环在复制起始区(IZ)定位中的作用。我们根据角点的存在(表明环的存在)和 CTCF 基序的取向来划分 TAD 和亚 TAD。我们发现,高效、早期复制的 IZ 定位于由高密度的发散和会聚取向 CTCF 基序锚定的相邻角点 TAD 之间的边界。相比之下,低效率的 IZ 定位于较弱的无点边界。在 G1 期间用 cohesin 介导的环挤压切除后,高效的 IZ 变得弥散和定位在具有复杂 CTCF 基序取向的边界处。此外,在 G1 敲低 cohesin 卸载因子 WAPL 会导致长距离环的获得,并且 IZ 在相同边界处的定位变窄。最后,特定边界的靶向缺失或插入分别导致局部复制时间的改变,与 IZ 的丢失或获得一致。我们的数据支持这样一种模型,即 cohesin 介导的环挤压和在一组特定的遗传编码 TAD 和亚 TAD 边界处的停滞是人类 S 期复制起始点位置的一个重要决定因素。