Research Team "Bacterial Opportunistic Pathogens and Environment" (BPOE), University of Lyon, UMR Ecologie Microbienne Lyon (LEM), Lyon, France.
Adipocyte. 2022 Dec;11(1):510-528. doi: 10.1080/21623945.2022.2107784.
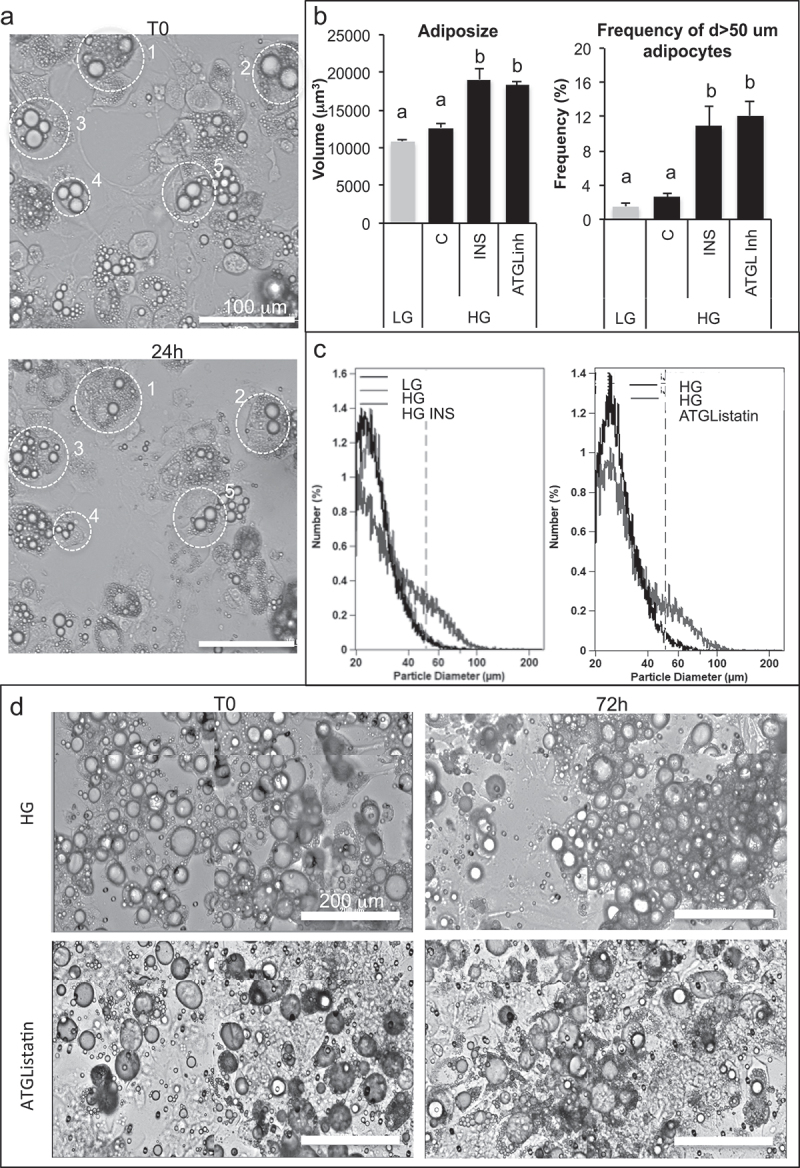
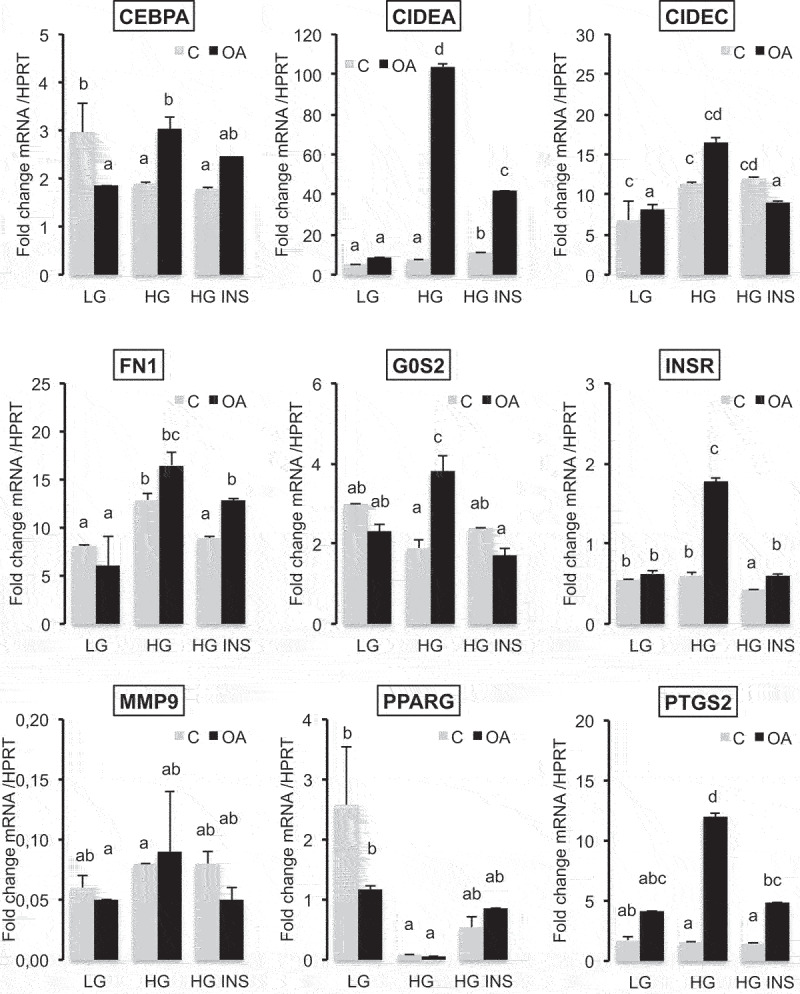
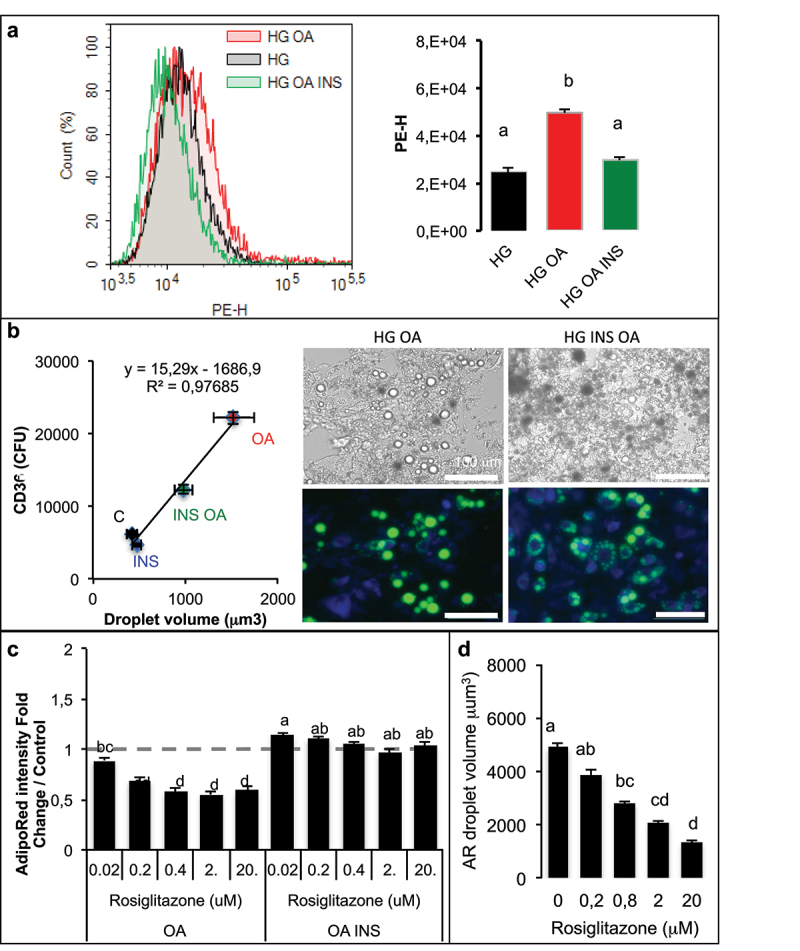
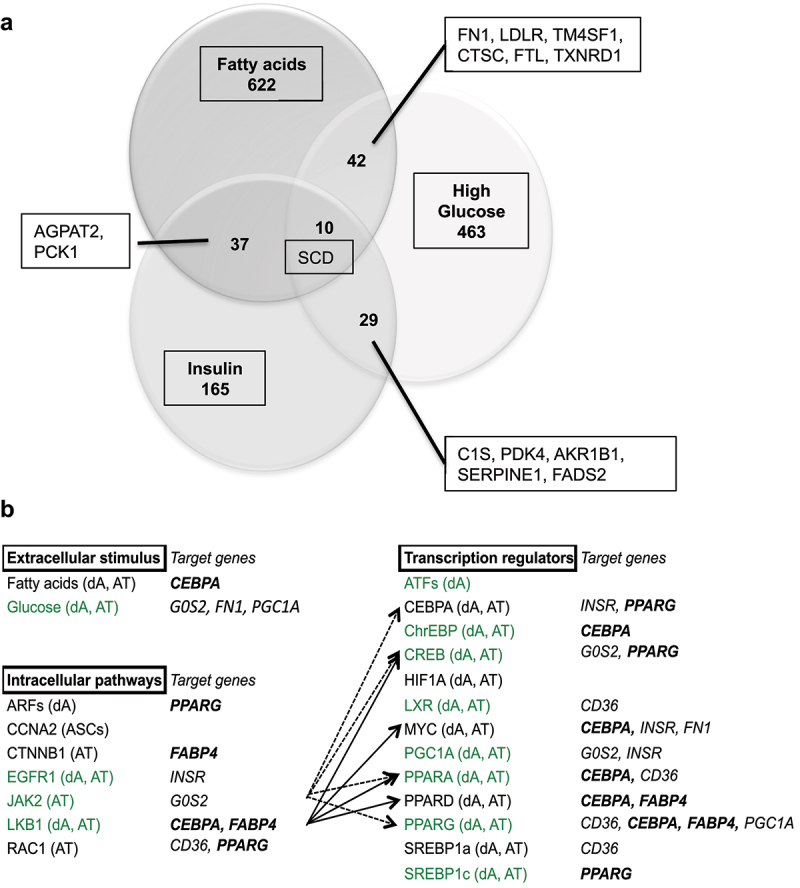
Metabolic disorders related to obesity are largely dependent on adipose tissue hypertrophy, which involves adipocyte hypertrophy and increased adipogenesis. Adiposize is regulated by lipid accumulation as a result of increased lipogenesis (mainly lipid uptake in mature adipocytes) and reduced lipolysis. Using realtime 2D cell culture analyses of lipid uptake, we show (1) that high glucose concentration (4.5 g/L) was required to accumulate oleic acid increasing lipid droplet size until unilocularization similar to mature adipocytes in few days, (2) oleic acid reduced ( gene transcription and (3) insulin counteracted oleic acid-induced increase of lipid droplet size. Although the lipolytic activity observed in high low glucose (1 g/L) conditions was not altered, insulin was found to inhibit oleic acid induced gene transcription required for lipid storage such as Cell Death Inducing DFFA Like Effectors (CIDEC) and G0 switch gene S2), possibly through PPARA activity. Although this signalling pathway requires more detailed investigation, the results point out the differential mechanisms involved in the pro-adipogenic effect of insulin in absence its protective effect on adiposity in presence of oleic acid uptake.: AICAR, 5-Aminoimidazole-4-carboxamide-1-D-ribofuranoside; AMPK, AMP-Activated protein kinase, ASCs, adipose stem cell; ATGL, adipose triglyceride lipase; BSA, Bovine serum albumin; CEBPA, CCAAT enhancer binding protein alpha; CIDEs, Cell Death Inducing DFFA Like Effectors; dA, differentiated adipocyte; DMEM, Dulbecco's Modified Eagle's Medium; FABPs, Fatty Acid Binding Proteins; FAT/CD36, Fatty acid translocase; FCS, Foetal calf serum; FN1, fibronectin 1; FFA, free fatty acid; G0S2, G0 switch gene S2; GLUTs, Glucose transporters; GPR120, G protein-coupled receptor 120; HG, high glucose; HSL, hormone sensitive lipase; INSR, insulin receptor; LG, low glucose; OA, oleic acid; PBS, Phosphate buffer saline; PPARs, Peroxisome-Proliferator Activated Receptors; PKA, Protein kinase cyclic AMP-dependent; PKG, Protein kinase cyclic GMP dependent; PTGS2, cytochrome oxidase 2; RTCA, realtime cell analysis; TG, triglyceride.
与肥胖相关的代谢紊乱在很大程度上依赖于脂肪组织肥大,其中涉及脂肪细胞肥大和脂肪生成增加。脂肪生成受到脂质积累的调节,这是由于脂肪生成增加(主要是成熟脂肪细胞中的脂质摄取)和脂肪分解减少所致。通过实时 2D 细胞培养分析脂质摄取,我们表明:
高葡萄糖浓度(4.5g/L)是积累油酸增加脂滴大小直至类似成熟脂肪细胞的单室化所必需的,需要几天时间;
油酸降低了基因转录;
胰岛素拮抗油酸诱导的脂滴大小增加。
虽然在高(1g/L)低葡萄糖条件下观察到的脂肪分解活性没有改变,但发现胰岛素抑制了脂肪储存所需的油酸诱导的基因转录,如细胞死亡诱导 DFFA 样效应物(CIDEC)和 G0 开关基因 S2),可能通过 PPARA 活性。虽然这种信号通路需要更详细的研究,但结果指出了胰岛素在缺乏其对油酸摄取时的促脂肪生成作用的不同机制:
AICAR,5-氨基咪唑-4-甲酰胺-1-D-核糖呋喃核苷酸;
AMPK,AMP 激活的蛋白激酶;
ASC,脂肪干细胞;
ATGL,脂肪甘油三酯脂肪酶;
BSA,牛血清白蛋白;
CEBPA,CCAAT 增强子结合蛋白 alpha;
CIDECs,细胞死亡诱导 DFFA 样效应物;
dA,分化的脂肪细胞;
DMEM,Dulbecco 的改良 Eagle 培养基;
FABPs,脂肪酸结合蛋白;
FAT/CD36,脂肪酸转运蛋白;
FCS,胎牛血清;
FN1,纤维连接蛋白 1;
FFA,游离脂肪酸;
G0S2,G0 开关基因 S2;
GLUTs,葡萄糖转运蛋白;
GPR120,G 蛋白偶联受体 120;
HG,高葡萄糖;
HSL,激素敏感脂肪酶;
INSR,胰岛素受体;
LG,低葡萄糖;
OA,油酸;
PBS,磷酸盐缓冲盐水;
PPARs,过氧化物酶体增殖物激活受体;
PKA,蛋白激酶 cAMP 依赖性;
PKG,蛋白激酶 cGMP 依赖性;
PTGS2,细胞色素氧化酶 2;
RTCA,实时细胞分析;
TG,甘油三酯。