Department of Pharmacology, University of Texas Southwestern Medical Center, Dallas, United States.
Westlake Laboratory of Life Sciences and Biomedicine, Hangzhou, China.
Elife. 2022 Aug 12;11:e77848. doi: 10.7554/eLife.77848.
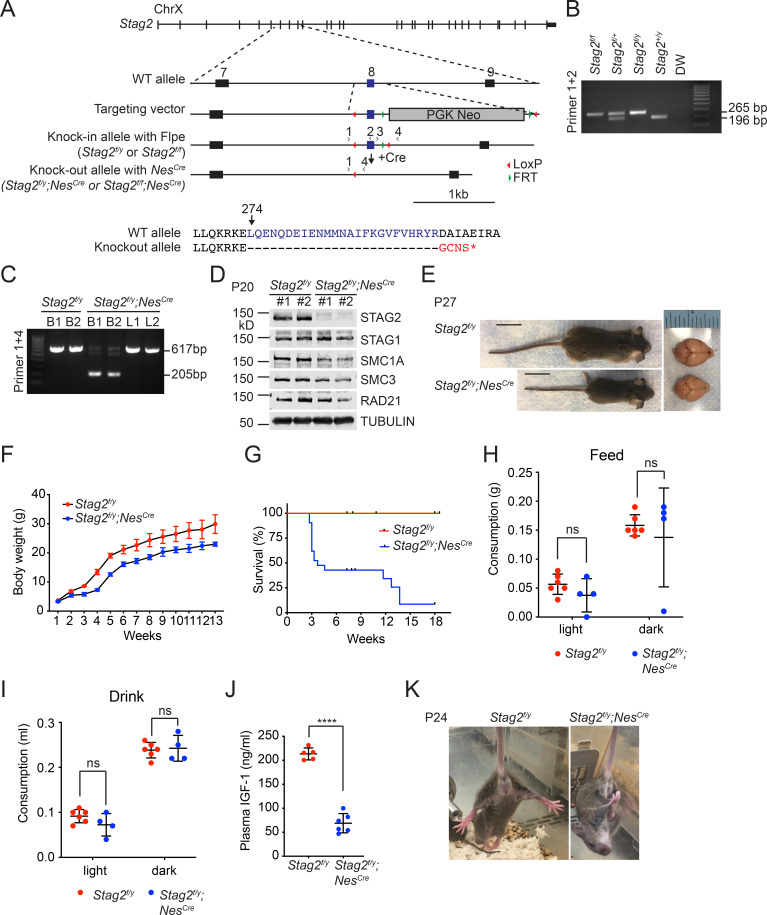
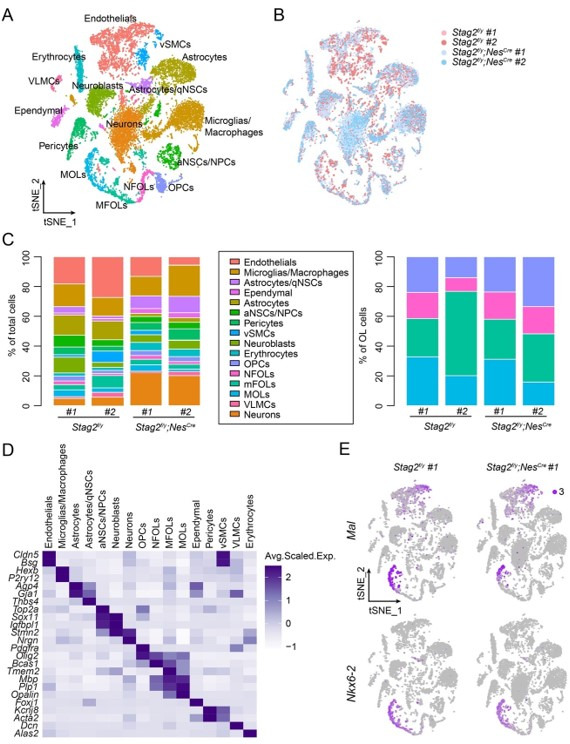
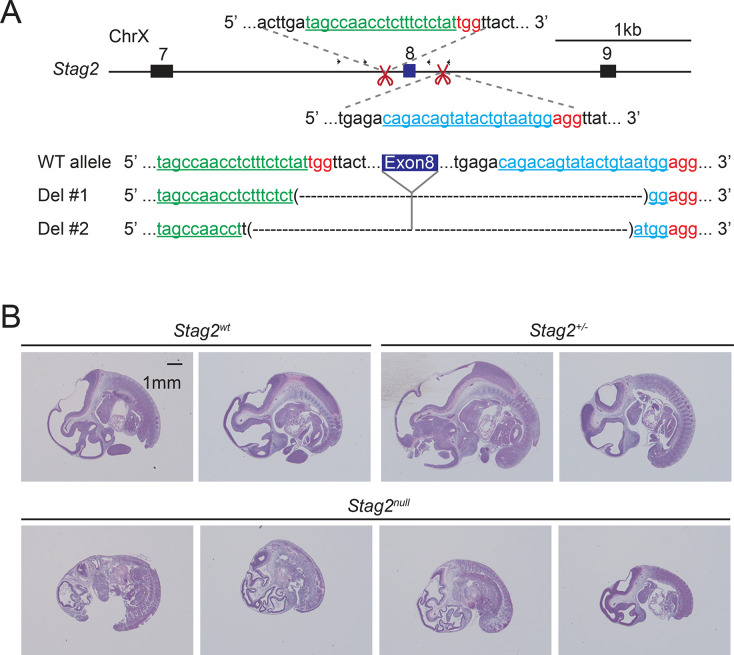
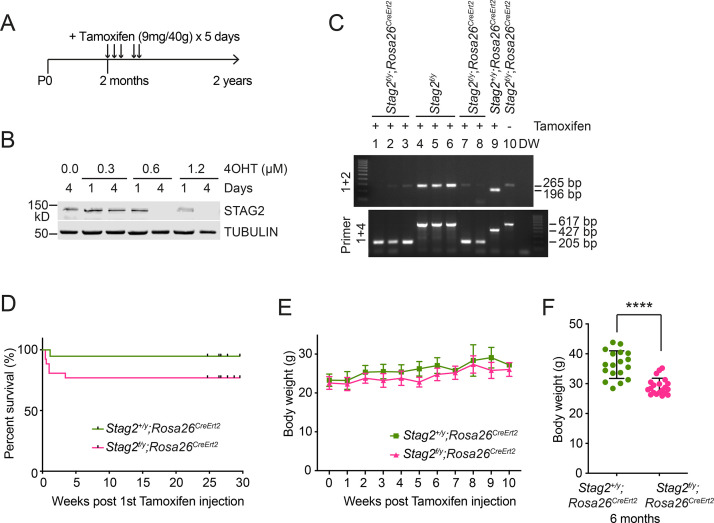
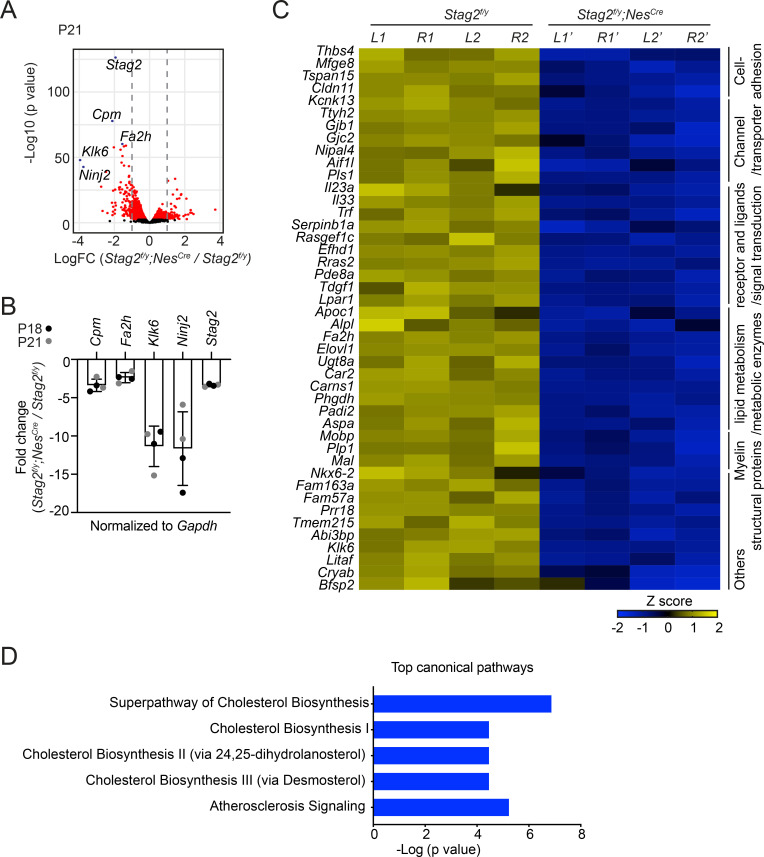
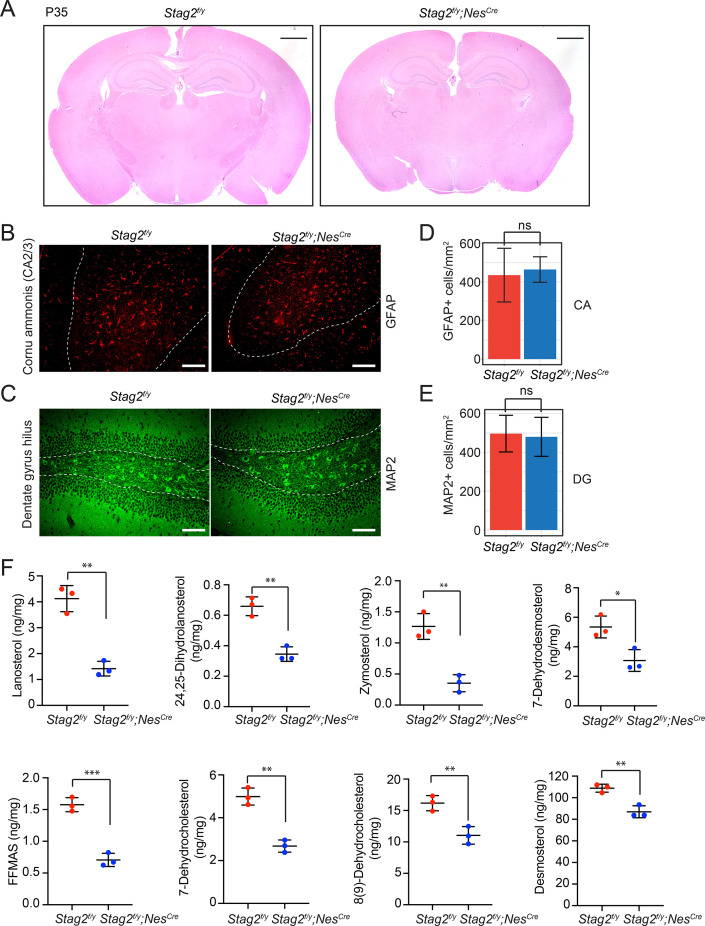
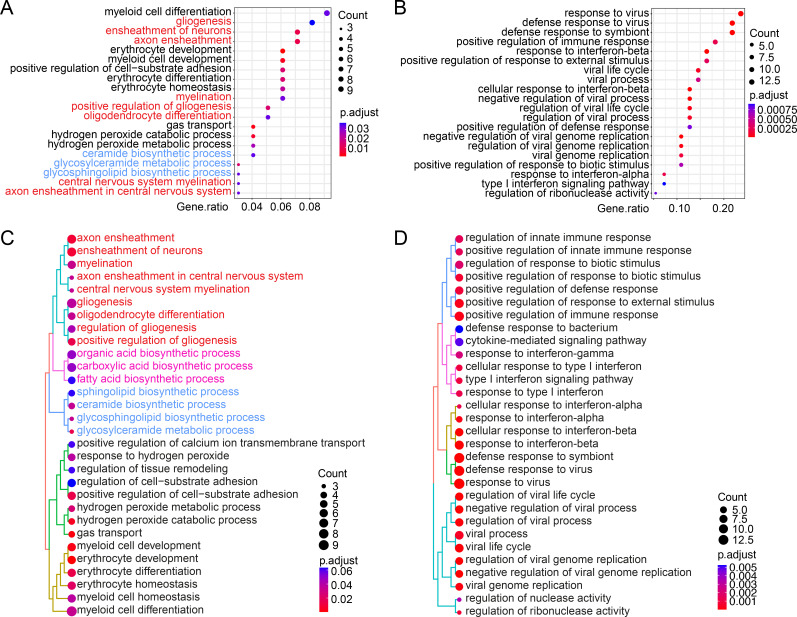
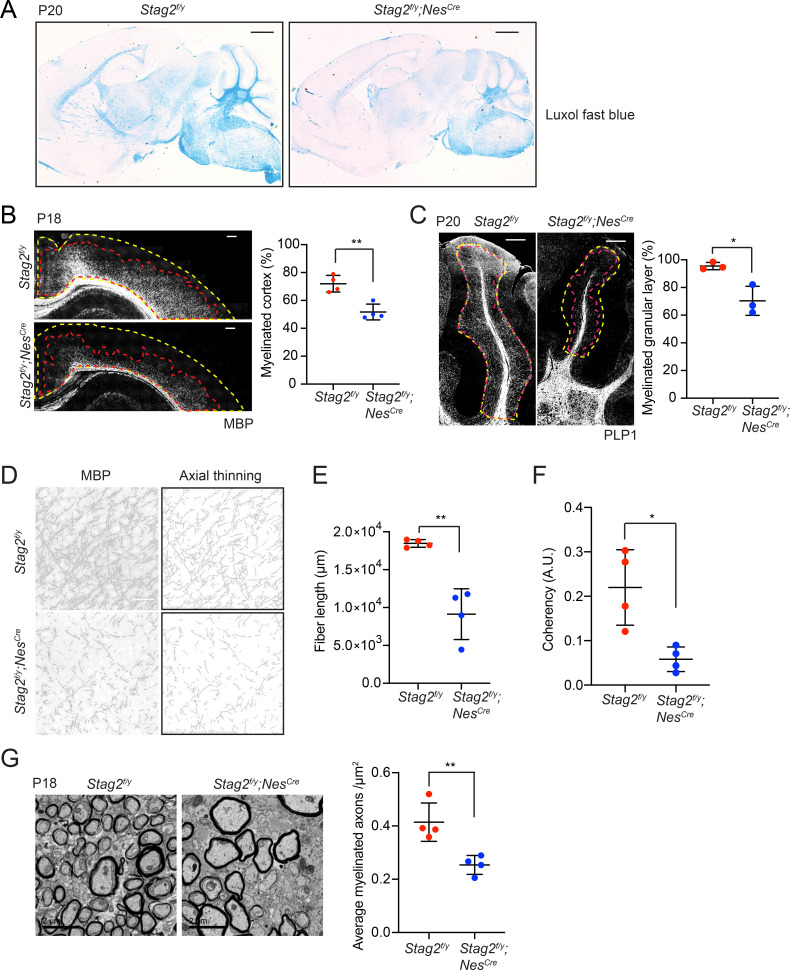
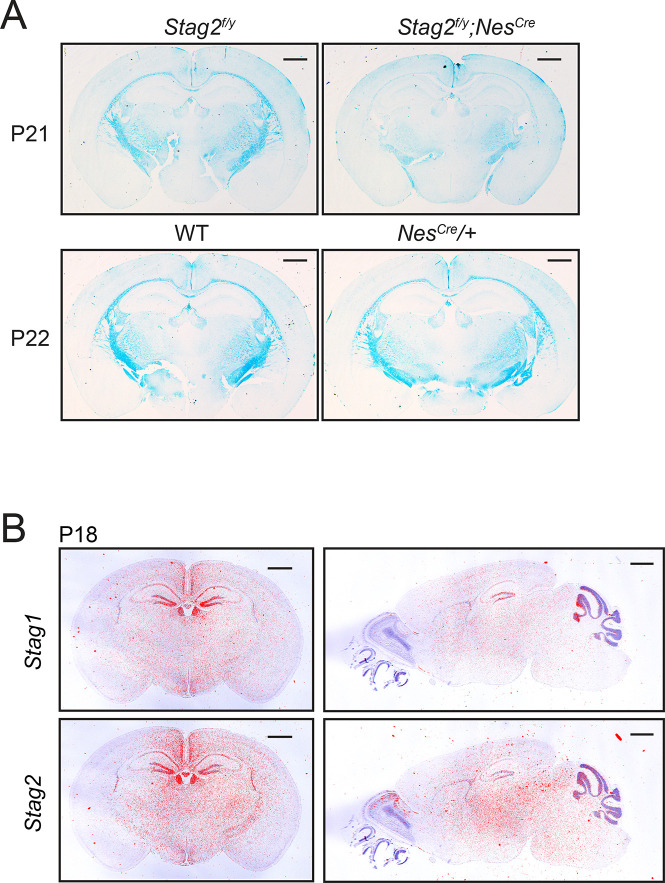
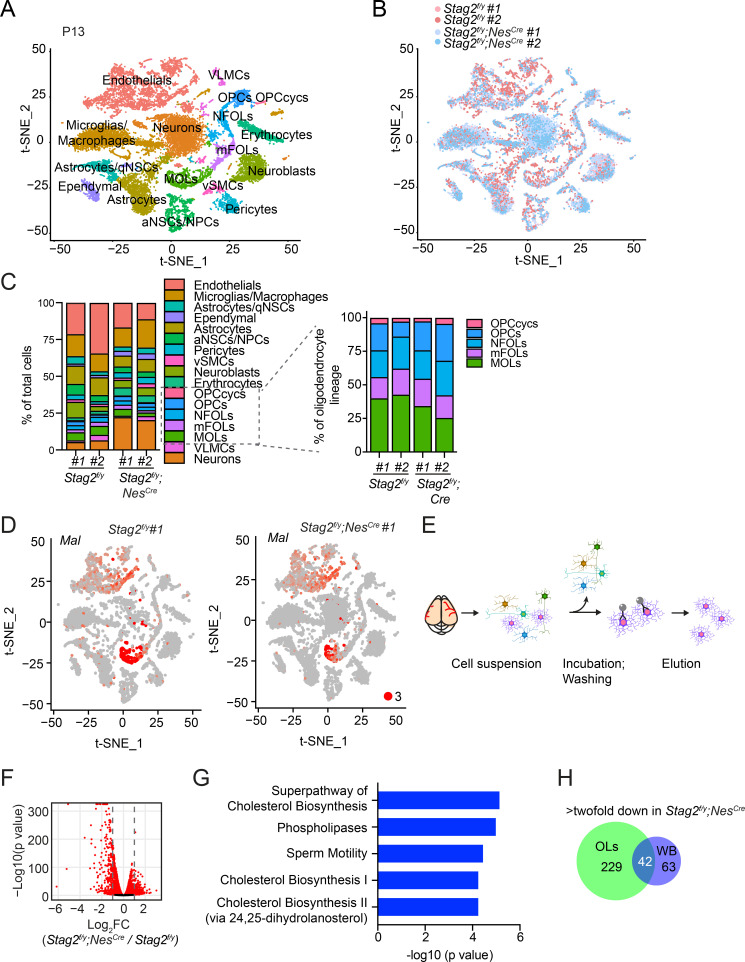
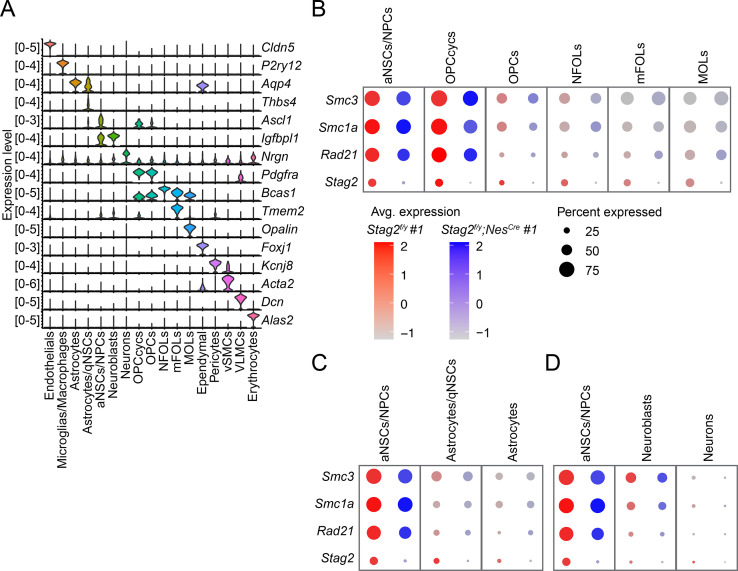
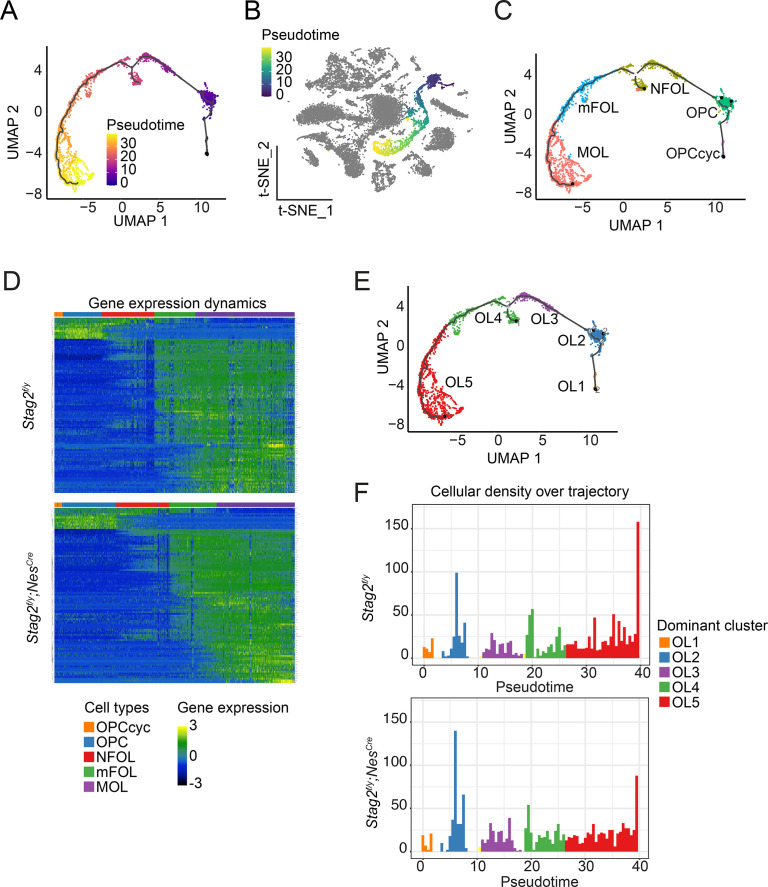
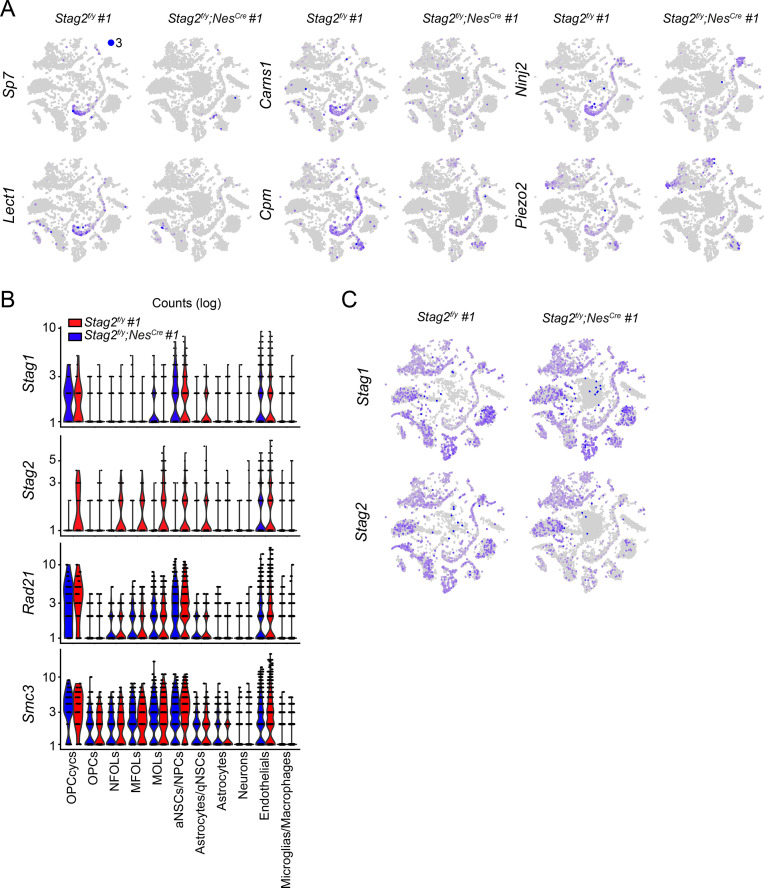
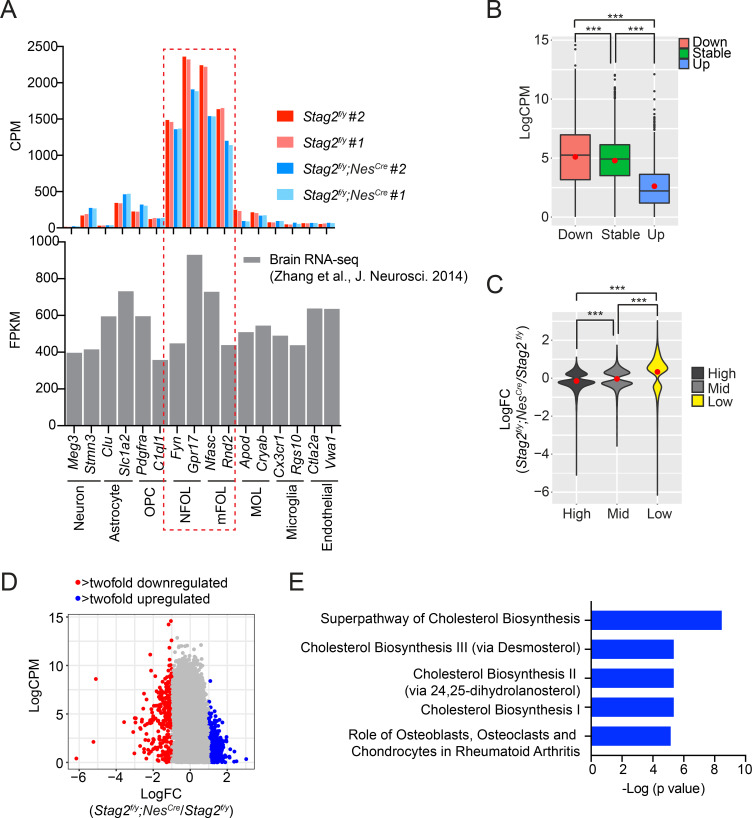
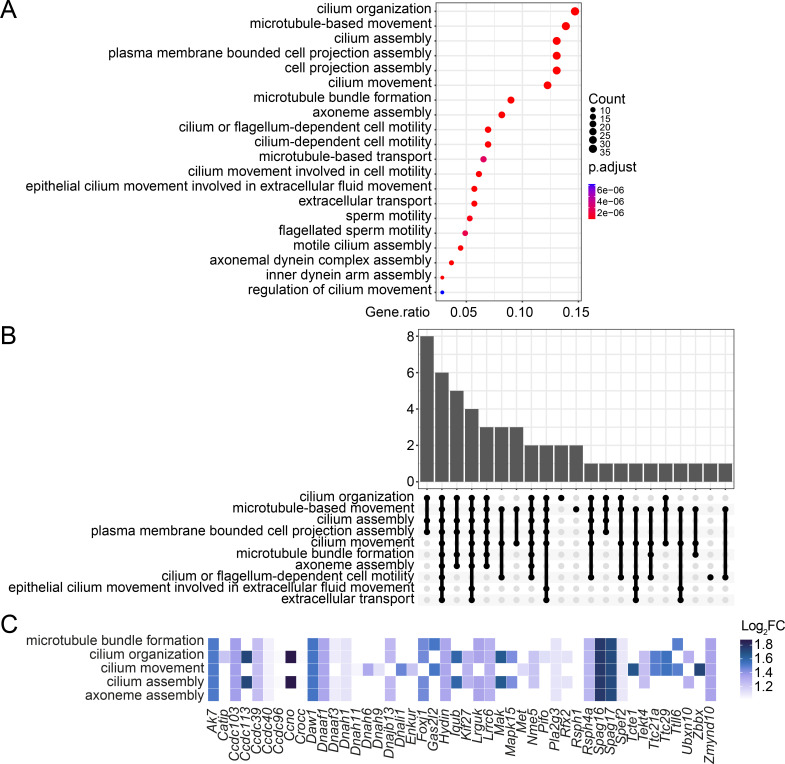
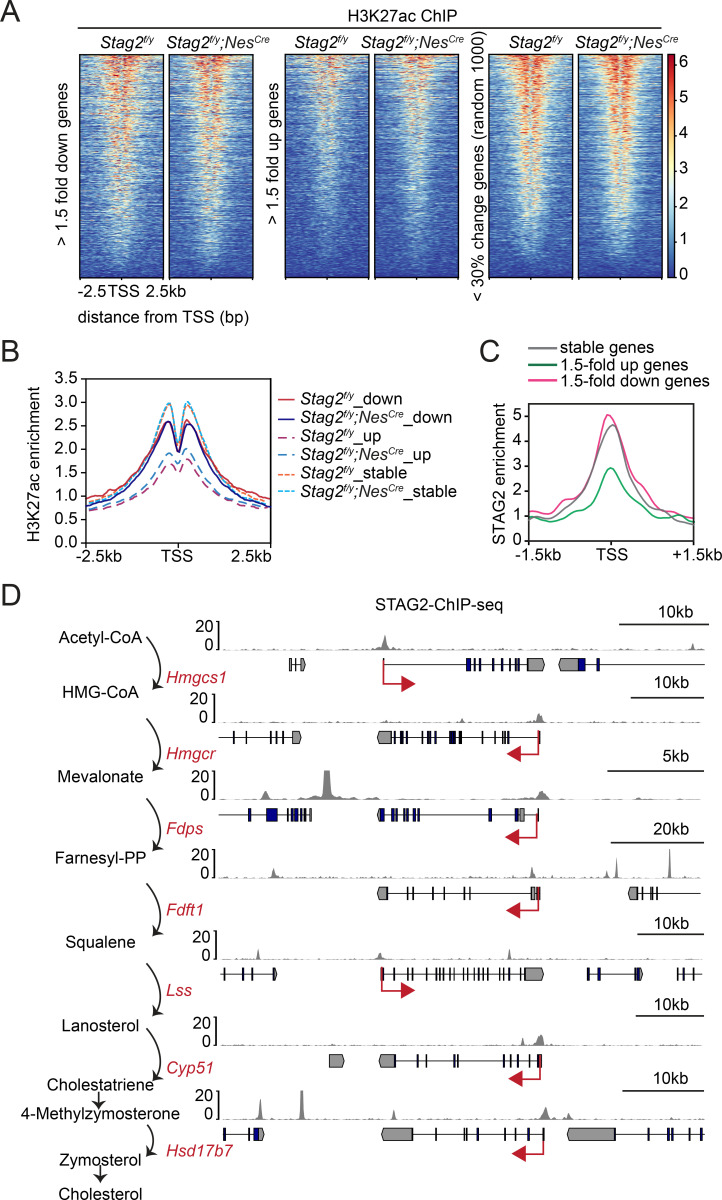
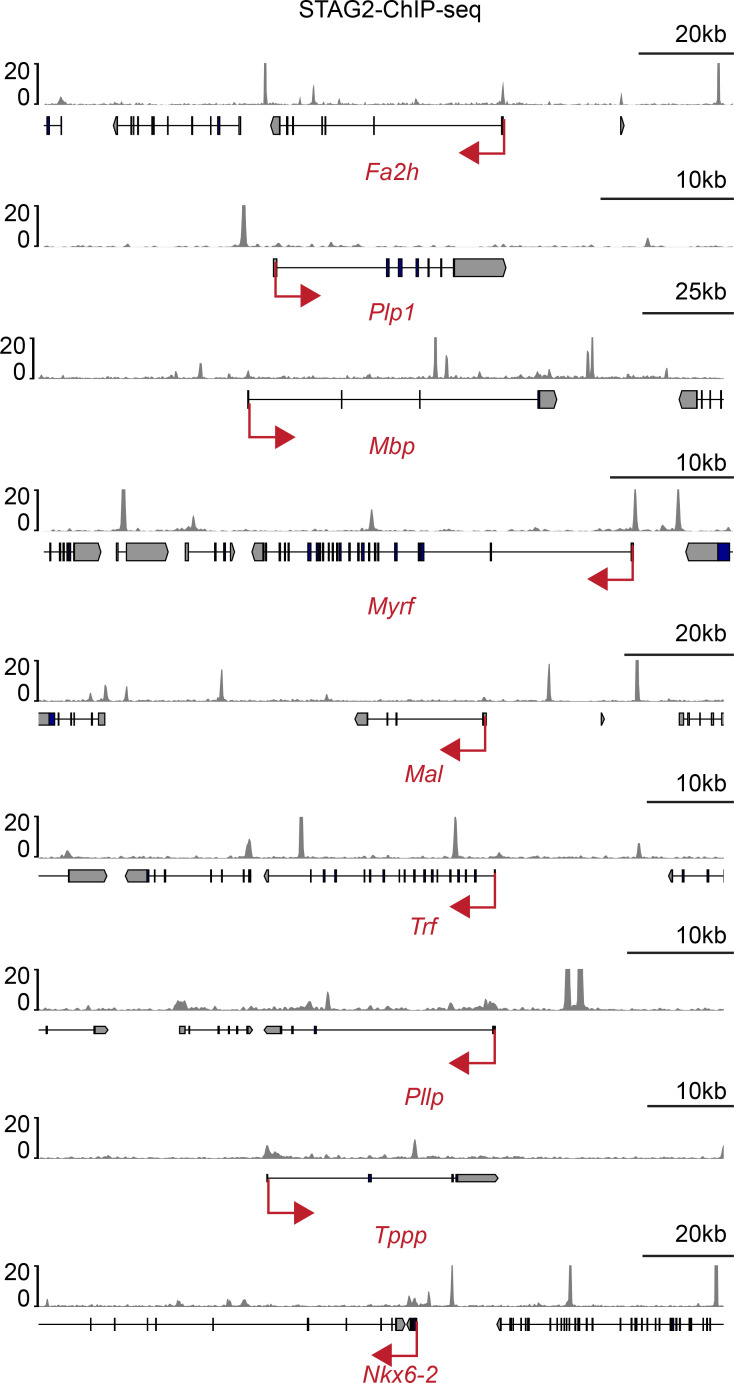
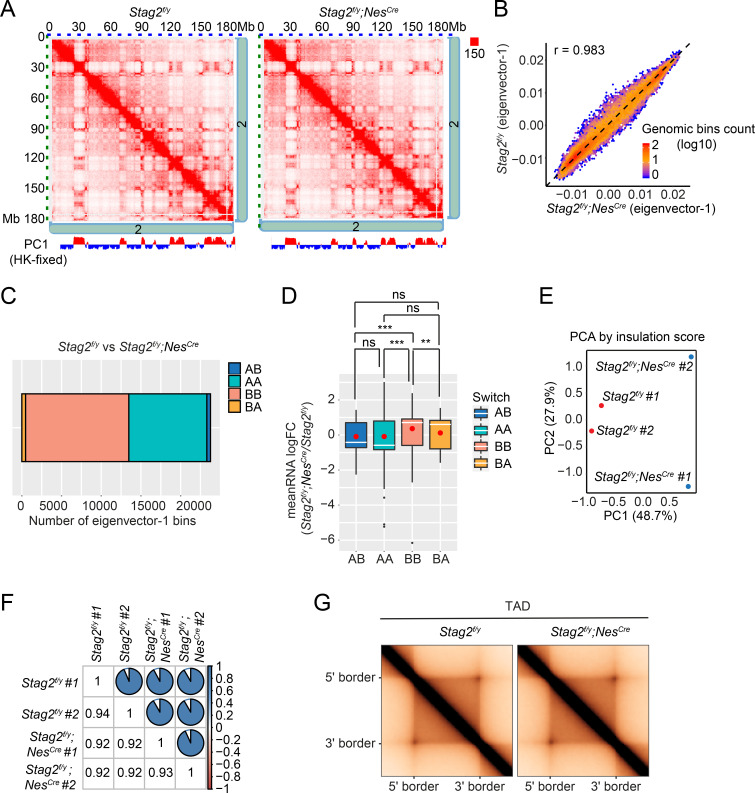
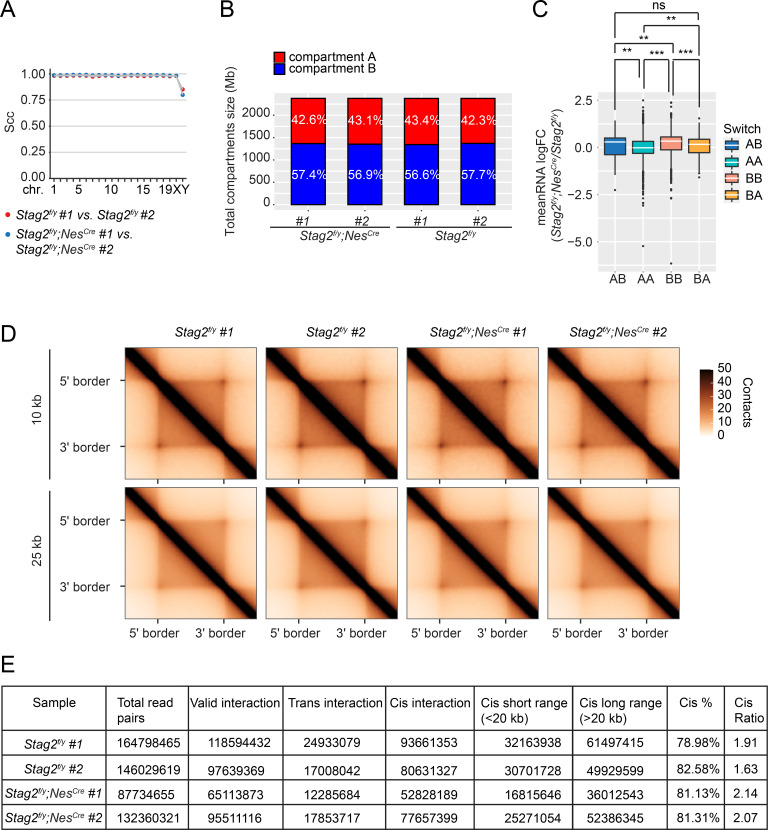
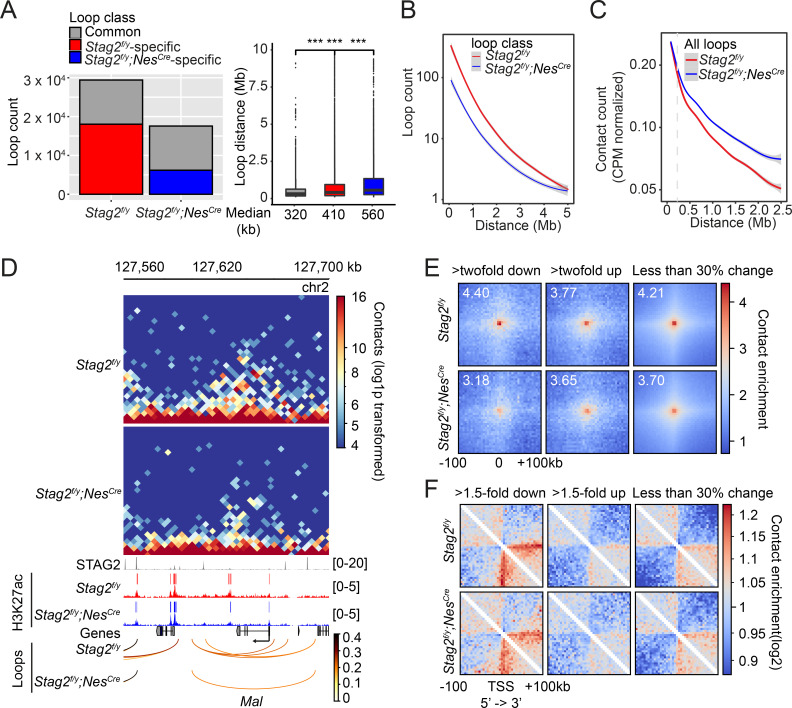
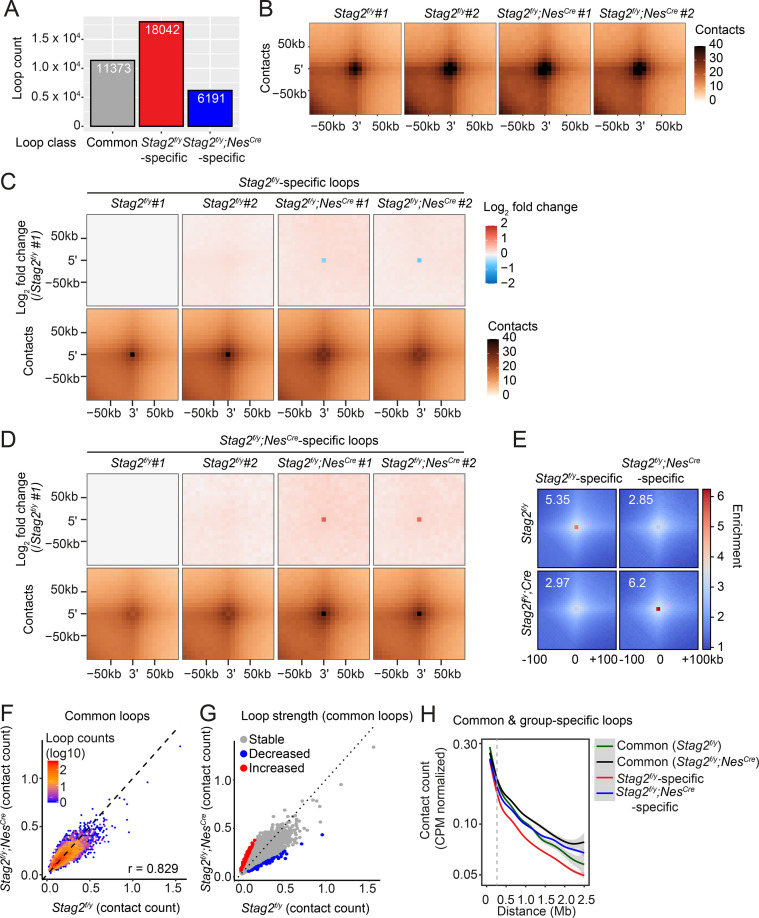
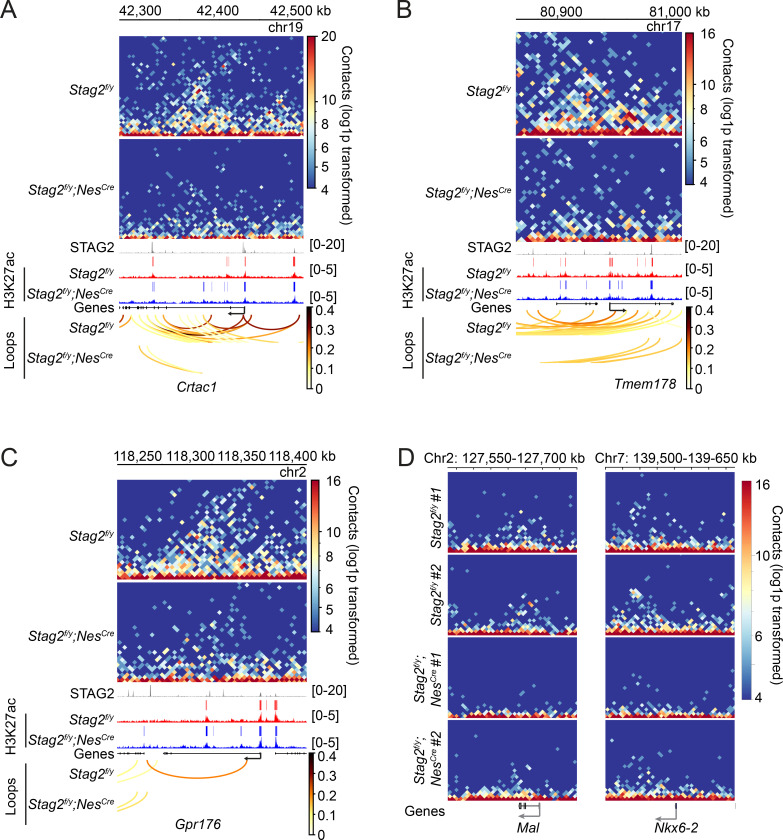
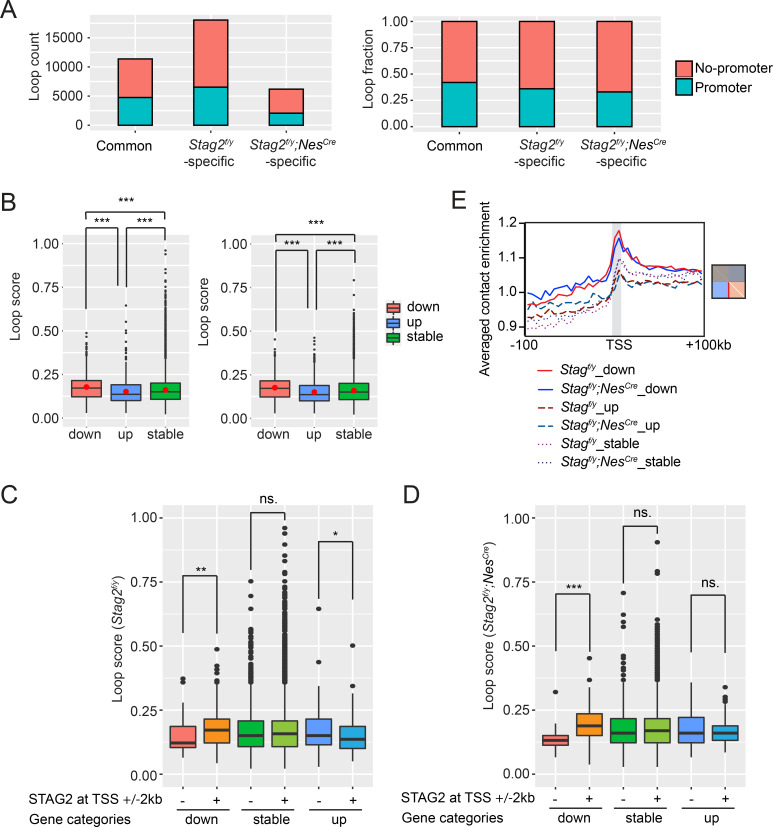
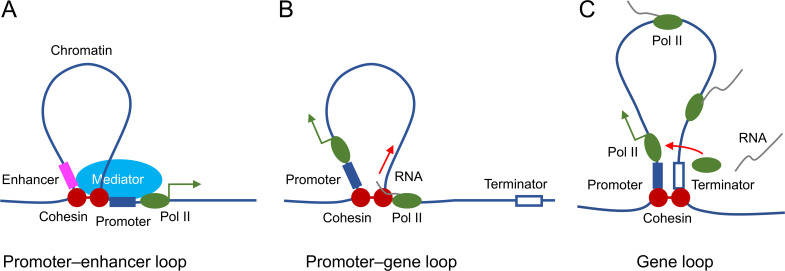
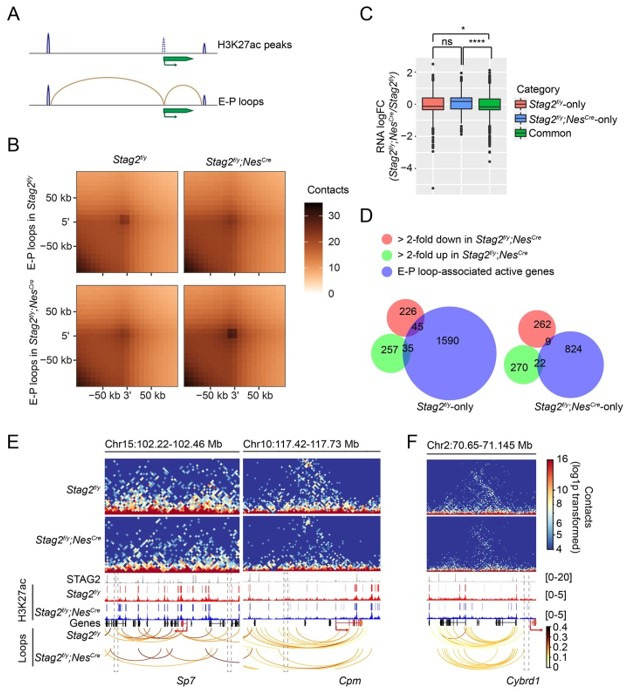
Cohesin folds chromosomes via DNA loop extrusion. Cohesin-mediated chromosome loops regulate transcription by shaping long-range enhancer-promoter interactions, among other mechanisms. Mutations of cohesin subunits and regulators cause human developmental diseases termed cohesinopathy. Vertebrate cohesin consists of SMC1, SMC3, RAD21, and either STAG1 or STAG2. To probe the physiological functions of cohesin, we created conditional knockout (cKO) mice with deleted in the nervous system. cKO mice exhibit growth retardation, neurological defects, and premature death, in part due to insufficient myelination of nerve fibers. cKO oligodendrocytes exhibit delayed maturation and downregulation of myelination-related genes. loss reduces promoter-anchored loops at downregulated genes in oligodendrocytes. Thus, STAG2-cohesin generates promoter-anchored loops at myelination-promoting genes to facilitate their transcription. Our study implicates defective myelination as a contributing factor to cohesinopathy and establishes oligodendrocytes as a relevant cell type to explore the mechanisms by which cohesin regulates transcription.
黏连蛋白通过 DNA 环伸出折叠染色体。黏连蛋白介导的染色体环通过塑造长程增强子-启动子相互作用等机制调节转录。黏连蛋白亚基和调节因子的突变会导致人类发育疾病,称为黏连蛋白病。脊椎动物黏连蛋白由 SMC1、SMC3、RAD21 和 STAG1 或 STAG2 组成。为了探究黏连蛋白的生理功能,我们在神经系统中创建了条件敲除(cKO)小鼠,该小鼠中 缺失。cKO 小鼠表现出生长迟缓、神经缺陷和过早死亡,部分原因是神经纤维髓鞘形成不足。cKO 少突胶质细胞表现出成熟延迟和髓鞘形成相关基因下调。缺失减少了少突胶质细胞中下调基因的启动子锚定环。因此,STAG2-黏连蛋白在促进髓鞘形成的基因上生成启动子锚定环,以促进其转录。我们的研究表明,髓鞘形成缺陷是黏连蛋白病的一个致病因素,并确立了少突胶质细胞作为一种相关细胞类型,以探索黏连蛋白调节转录的机制。