Cancer Biology and Genetics Program, Memorial Sloan Kettering Cancer Center, New York, NY, USA.
Department of Pharmacology, University of North Carolina at Chapel Hill School of Medicine, Chapel Hill, NC, USA.
Nature. 2022 Aug;608(7924):795-802. doi: 10.1038/s41586-022-05082-5. Epub 2022 Aug 17.
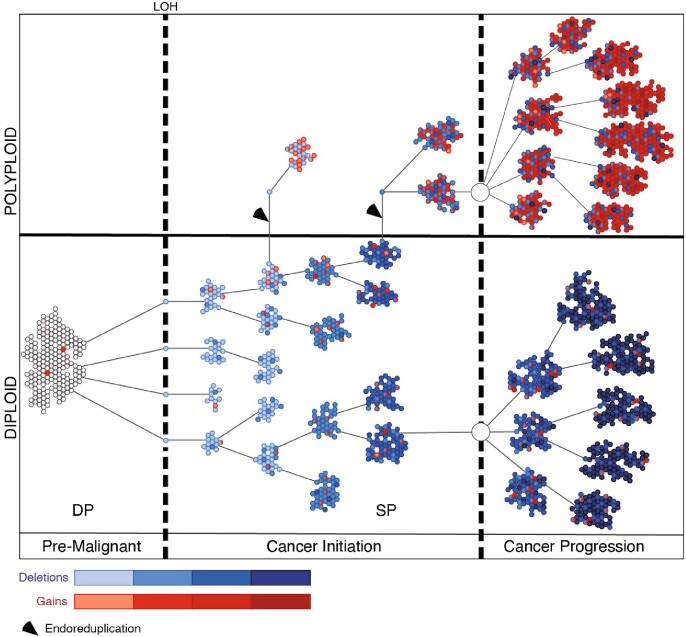
Although p53 inactivation promotes genomic instability and presents a route to malignancy for more than half of all human cancers, the patterns through which heterogenous TP53 (encoding human p53) mutant genomes emerge and influence tumorigenesis remain poorly understood. Here, in a mouse model of pancreatic ductal adenocarcinoma that reports sporadic p53 loss of heterozygosity before cancer onset, we find that malignant properties enabled by p53 inactivation are acquired through a predictable pattern of genome evolution. Single-cell sequencing and in situ genotyping of cells from the point of p53 inactivation through progression to frank cancer reveal that this deterministic behaviour involves four sequential phases-Trp53 (encoding mouse p53) loss of heterozygosity, accumulation of deletions, genome doubling, and the emergence of gains and amplifications-each associated with specific histological stages across the premalignant and malignant spectrum. Despite rampant heterogeneity, the deletion events that follow p53 inactivation target functionally relevant pathways that can shape genomic evolution and remain fixed as homogenous events in diverse malignant populations. Thus, loss of p53-the 'guardian of the genome'-is not merely a gateway to genetic chaos but, rather, can enable deterministic patterns of genome evolution that may point to new strategies for the treatment of TP53-mutant tumours.
尽管 p53 失活会促进基因组不稳定,并为超过一半的人类癌症提供恶性转化途径,但异质性 TP53(编码人类 p53)突变基因组出现并影响肿瘤发生的模式仍知之甚少。在这里,在一种报告在癌症发生前散发性 p53 杂合性丢失的胰腺导管腺癌小鼠模型中,我们发现 p53 失活所赋予的恶性特性是通过可预测的基因组进化模式获得的。从 p53 失活到进展为明确癌症的细胞的单细胞测序和原位基因分型揭示了这种确定性行为涉及四个连续阶段-Trp53(编码小鼠 p53)杂合性丢失、缺失积累、基因组加倍以及增益和扩增的出现-每个阶段都与癌前和恶性谱中的特定组织学阶段相关。尽管存在广泛的异质性,但紧随 p53 失活之后的缺失事件靶向功能相关途径,这些途径可以塑造基因组进化,并在不同的恶性群体中作为同质事件固定下来。因此,p53(“基因组的守护者”)的丧失不仅是通向遗传混乱的门户,而且可以实现基因组进化的确定性模式,这可能为治疗 TP53 突变肿瘤指明新的策略。