Oncology Research, Amgen Research, Thousand Oaks, CA 91320.
Department of Cellular and Molecular Pharmacology, University of California, San Francisco, CA 94158.
Proc Natl Acad Sci U S A. 2022 Sep 6;119(36):e2205608119. doi: 10.1073/pnas.2205608119. Epub 2022 Aug 29.
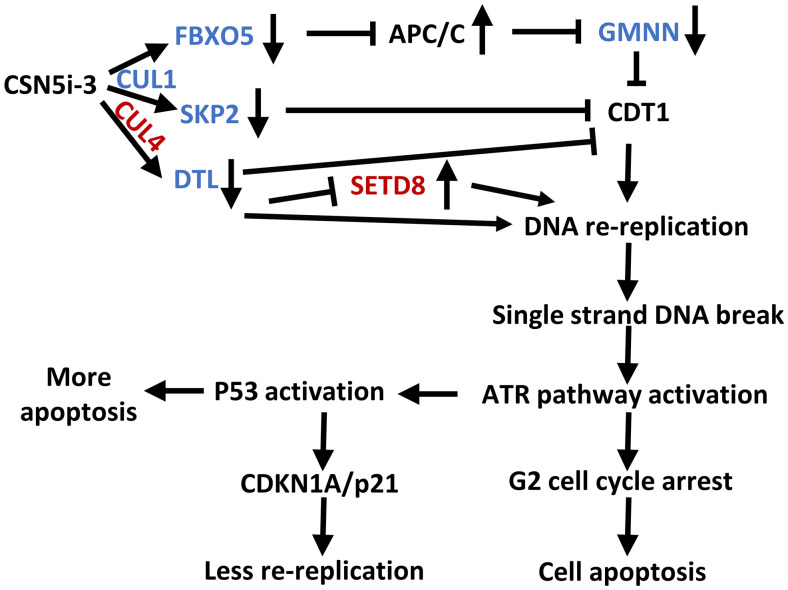
Cop9 signalosome (CSN) regulates the function of cullin-RING E3 ubiquitin ligases (CRLs) by deconjugating the ubiquitin-like protein NEDD8 from the cullin subunit. To understand the physiological impact of CSN function on the CRL network and cell proliferation, we combined quantitative mass spectrometry and genome-wide CRISPR interference (CRISPRi) and CRISPR activation (CRISPRa) screens to identify factors that modulate cell viability upon inhibition of CSN by the small molecule CSN5i-3. CRL components and regulators strongly modulated the antiproliferative effects of CSN5i-3, and in addition we found two pathways involved in genome integrity, SCF-APC/C-GMNN and CUL4-SETD8, that contribute substantially to the toxicity of CSN inhibition. Our data highlight the importance of CSN-mediated NEDD8 deconjugation and adaptive exchange of CRL substrate receptors in sustaining CRL function and suggest approaches for leveraging CSN inhibition for the treatment of cancer.
COP9 信号小体 (CSN) 通过从连接酶亚基上除去类泛素蛋白 NEDD8 来调节 Cullin-RING E3 连接酶 (CRL) 的功能。为了了解 CSN 功能对 CRL 网络和细胞增殖的生理影响,我们结合定量质谱和全基因组 CRISPR 干扰 (CRISPRi) 和 CRISPR 激活 (CRISPRa) 筛选,以鉴定在小分子 CSN5i-3 抑制 CSN 时调节细胞活力的因素。CRL 成分和调节剂强烈调节 CSN5i-3 的抗增殖作用,此外,我们还发现了两个涉及基因组完整性的途径,SCF-APC/C-GMNN 和 CUL4-SETD8,它们对 CSN 抑制的毒性有很大贡献。我们的数据强调了 CSN 介导的 NEDD8 去共轭和 CRL 底物受体的适应性交换在维持 CRL 功能中的重要性,并提出了利用 CSN 抑制治疗癌症的方法。