Department of Molecular Machines and Signaling, Max Planck Institute of Biochemistry, Martinsried, Germany.
Carle Illinois College of Medicine, Champaign, IL, USA.
Nature. 2021 Feb;590(7847):671-676. doi: 10.1038/s41586-021-03197-9. Epub 2021 Feb 3.
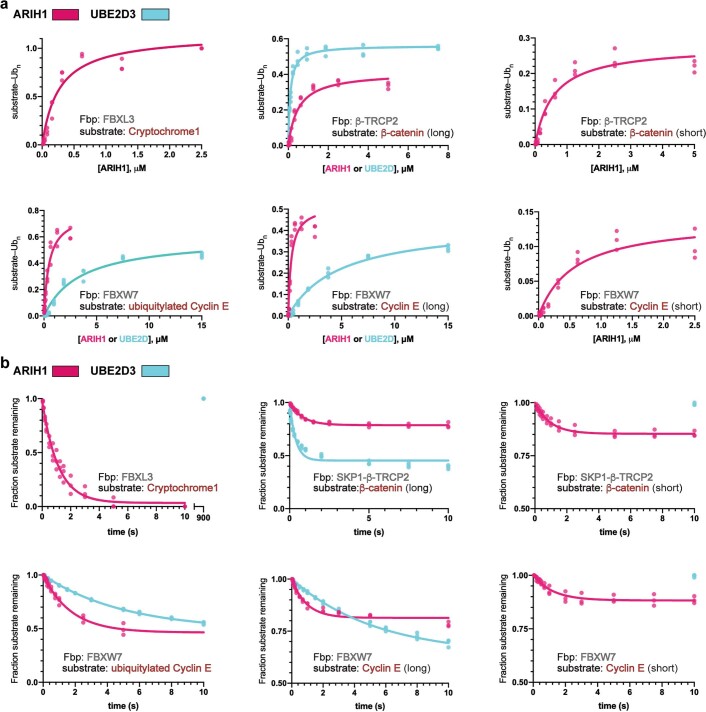
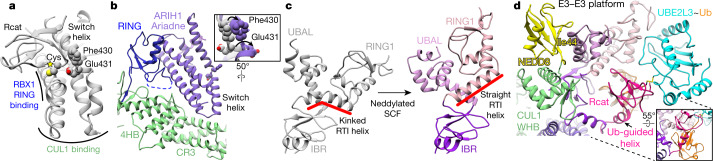
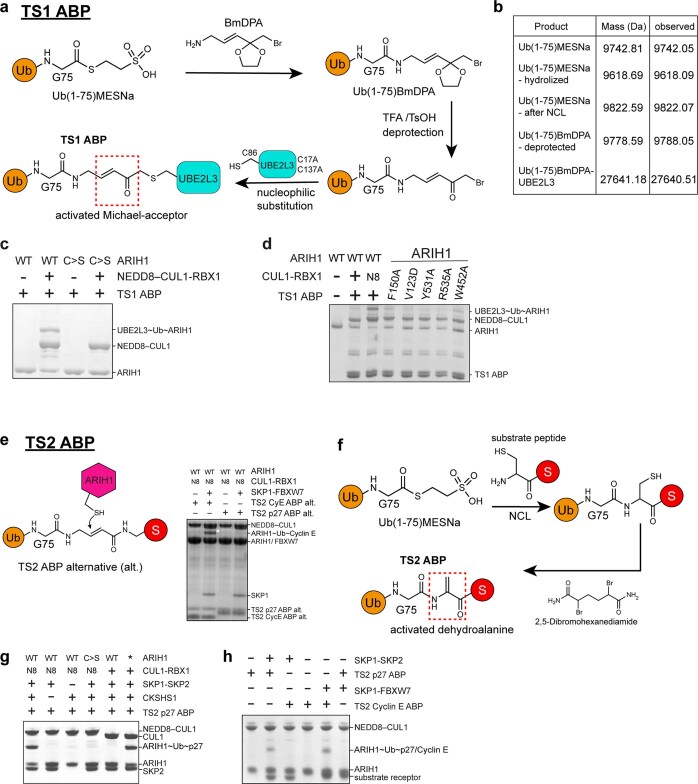
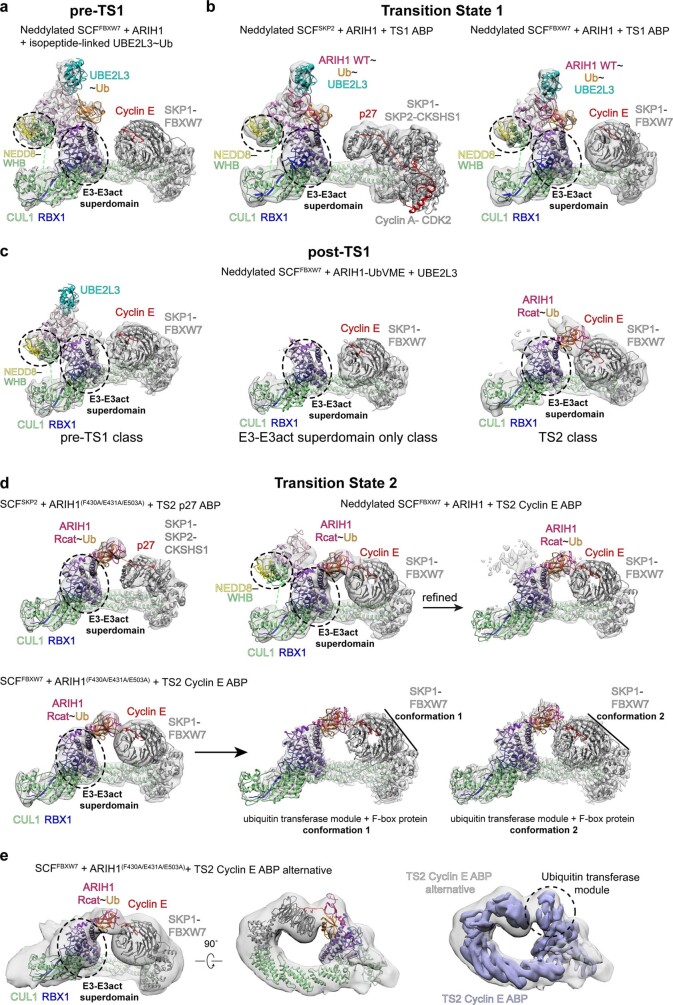
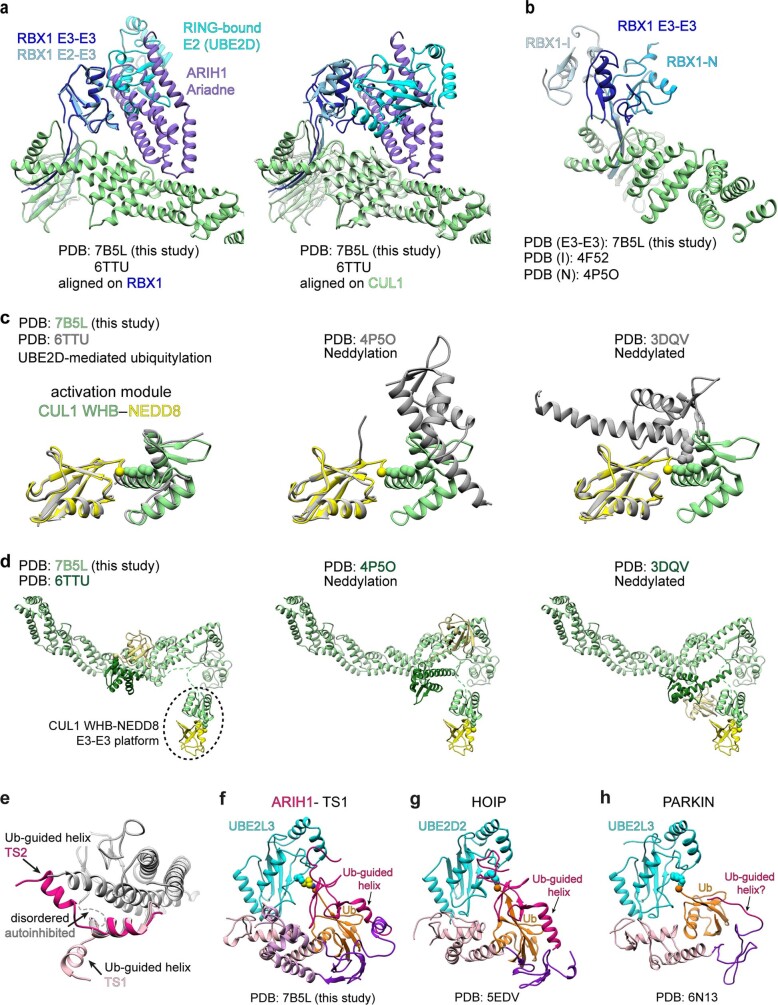
E3 ligases are typically classified by hallmark domains such as RING and RBR, which are thought to specify unique catalytic mechanisms of ubiquitin transfer to recruited substrates. However, rather than functioning individually, many neddylated cullin-RING E3 ligases (CRLs) and RBR-type E3 ligases in the ARIH family-which together account for nearly half of all ubiquitin ligases in humans-form E3-E3 super-assemblies. Here, by studying CRLs in the SKP1-CUL1-F-box (SCF) family, we show how neddylated SCF ligases and ARIH1 (an RBR-type E3 ligase) co-evolved to ubiquitylate diverse substrates presented on various F-box proteins. We developed activity-based chemical probes that enabled cryo-electron microscopy visualization of steps in E3-E3 ubiquitylation, initiating with ubiquitin linked to the E2 enzyme UBE2L3, then transferred to the catalytic cysteine of ARIH1, and culminating in ubiquitin linkage to a substrate bound to the SCF E3 ligase. The E3-E3 mechanism places the ubiquitin-linked active site of ARIH1 adjacent to substrates bound to F-box proteins (for example, substrates with folded structures or limited length) that are incompatible with previously described conventional RING E3-only mechanisms. The versatile E3-E3 super-assembly may therefore underlie widespread ubiquitylation.
E3 连接酶通常根据特征结构域进行分类,例如 RING 和 RBR,这些结构域被认为指定了泛素向募集底物转移的独特催化机制。然而,许多被 neddylation 的 cullin-RING E3 连接酶 (CRLs) 和 RBR 型 E3 连接酶家族中的 ARIH——它们共同构成了人类中几乎所有泛素连接酶的近一半——形成 E3-E3 超级组装体。在这里,通过研究 SKP1-CUL1-F-box (SCF) 家族中的 CRLs,我们展示了 neddylated SCF 连接酶和 ARIH1(一种 RBR 型 E3 连接酶)如何共同进化以泛素化各种不同的底物,这些底物呈现在各种 F-box 蛋白上。我们开发了基于活性的化学探针,使 cryo-electron microscopy 能够可视化 E3-E3 泛素化的步骤,从与 E2 酶 UBE2L3 连接的泛素开始,然后转移到 ARIH1 的催化半胱氨酸,最后与结合到 SCF E3 连接酶的底物连接。E3-E3 机制将 ARIH1 的泛素连接活性位点置于与 F-box 蛋白结合的底物(例如,具有折叠结构或有限长度的底物)附近,这些底物与先前描述的传统 RING E3 仅有的机制不兼容。因此,多功能的 E3-E3 超级组装体可能是广泛泛素化的基础。