Department of Molecular Machines and Signaling, Max Planck Institute of Biochemistry, Martinsried, Germany.
School of Pharmacy, University of Waterloo, Waterloo, Ontario, Canada.
Nat Chem Biol. 2023 Dec;19(12):1513-1523. doi: 10.1038/s41589-023-01392-5. Epub 2023 Aug 31.
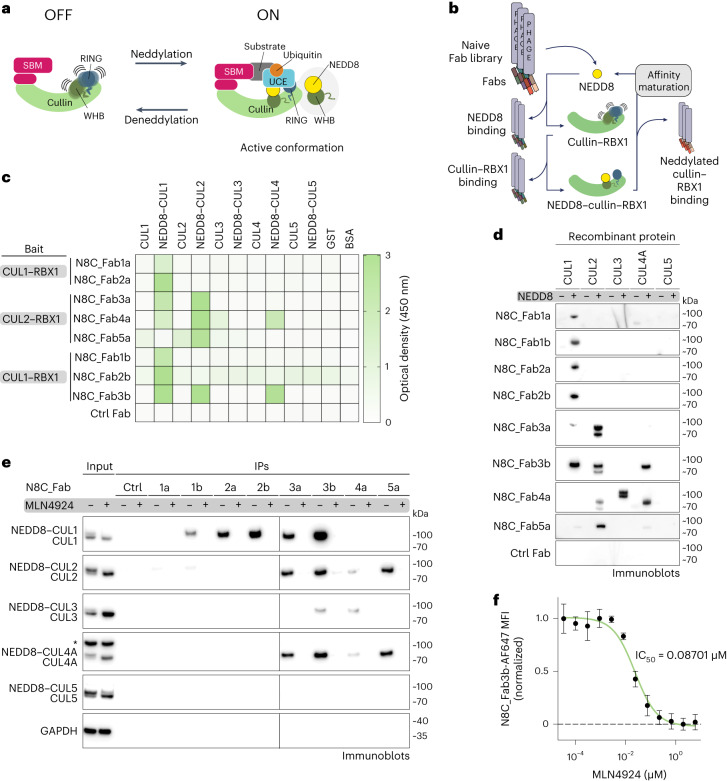
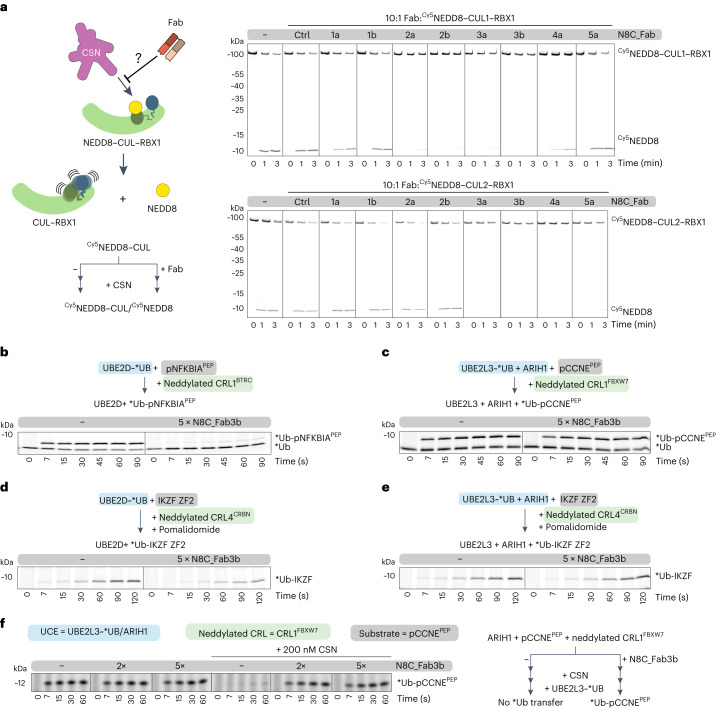
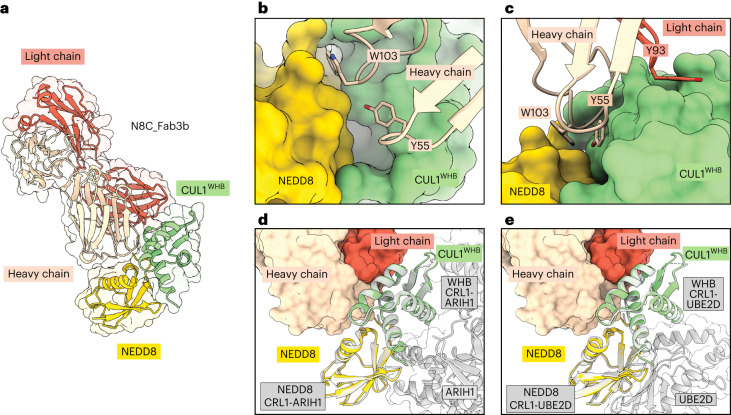
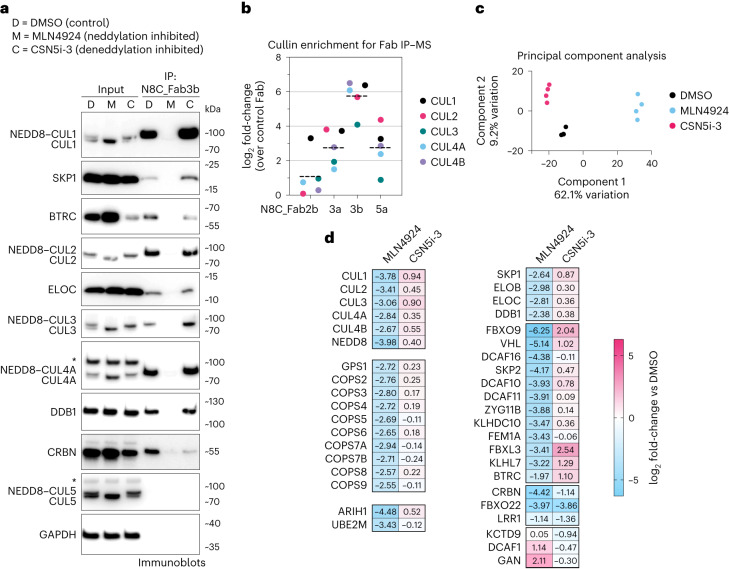
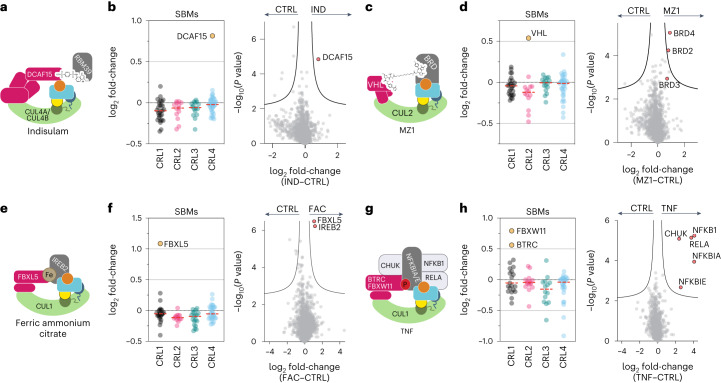
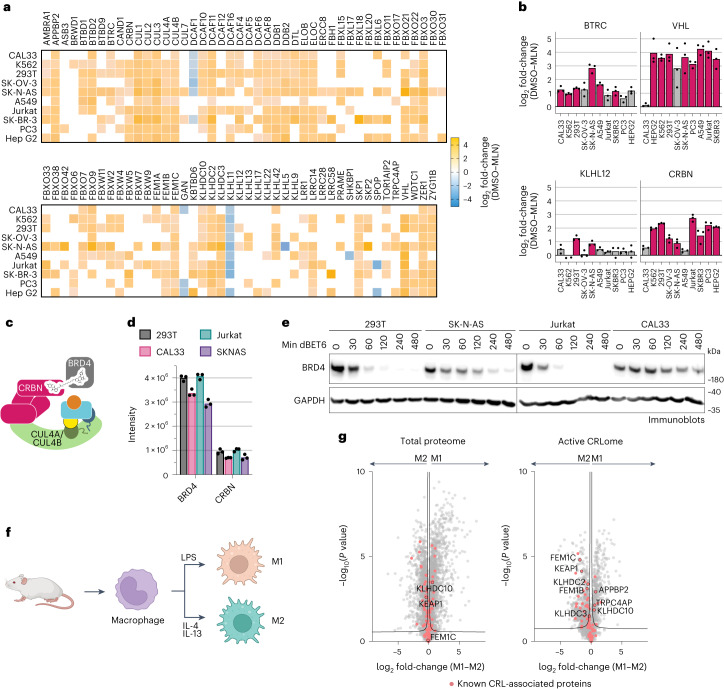
The cullin-RING ubiquitin ligase (CRL) network comprises over 300 unique complexes that switch from inactive to activated conformations upon site-specific cullin modification by the ubiquitin-like protein NEDD8. Assessing cellular repertoires of activated CRL complexes is critical for understanding eukaryotic regulation. However, probes surveying networks controlled by site-specific ubiquitin-like protein modifications are lacking. We developed a synthetic antibody recognizing the active conformation of NEDD8-linked cullins. Implementing the probe to profile cellular networks of activated CUL1-, CUL2-, CUL3- and CUL4-containing E3s revealed the complexes responding to stimuli. Profiling several cell types showed their baseline neddylated CRL repertoires vary, and prime efficiency of targeted protein degradation. Our probe also unveiled differential rewiring of CRL networks across distinct primary cell activation pathways. Thus, conformation-specific probes can permit nonenzymatic activity-based profiling across a system of numerous multiprotein complexes, which in the case of neddylated CRLs reveals widespread regulation and could facilitate the development of degrader drugs.
Cullin-RING 泛素连接酶 (CRL) 网络由 300 多种独特的复合物组成,这些复合物在特异性 Cullin 修饰后,通过类泛素蛋白 NEDD8 从非活性构象转变为激活构象。评估细胞中激活的 CRL 复合物的组成对于理解真核生物的调控至关重要。然而,目前缺乏用于探测受特定泛素样蛋白修饰调控的网络的探针。我们开发了一种能够识别 NEDD8 连接的 Cullin 激活构象的合成抗体。利用该探针对包含 CUL1、CUL2、CUL3 和 CUL4 的 E3 的细胞内网络进行分析,揭示了对刺激作出响应的复合物。对几种细胞类型进行分析表明,它们的基础 NEDDYLATED CRL 组成不同,靶向蛋白降解的效率也不同。我们的探针还揭示了不同的原代细胞激活途径中 CRL 网络的差异重排。因此,构象特异性探针可以实现对大量多蛋白复合物系统的非酶活性的基于活性的分析,对于 NEDDYLATED CRL 来说,这种方法揭示了广泛的调控作用,并可能有助于降解剂药物的开发。