Division of Biomedical Sciences, Warwick Medical School, University of Warwick, Coventry, United Kingdom.
Elife. 2022 Sep 6;11:e81755. doi: 10.7554/eLife.81755.
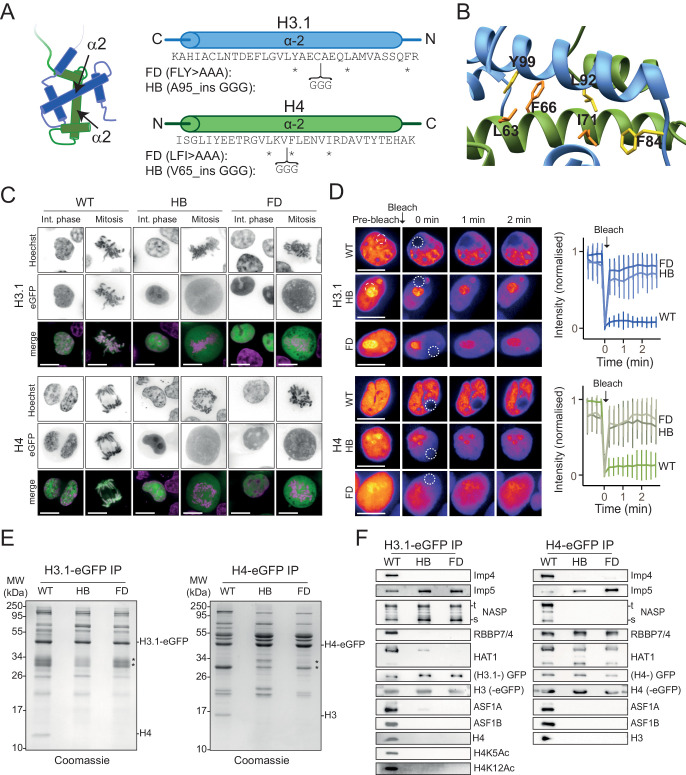
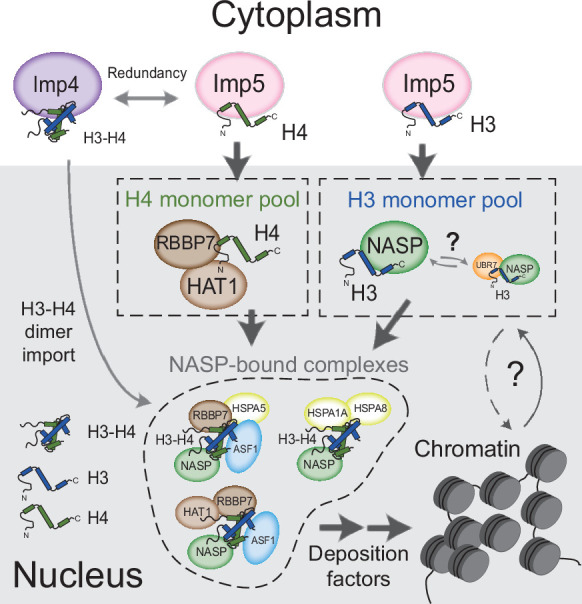
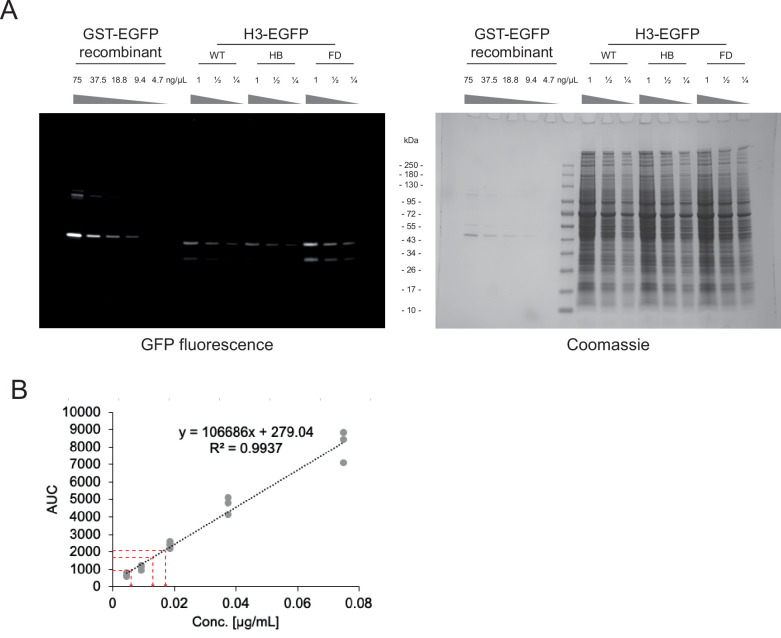
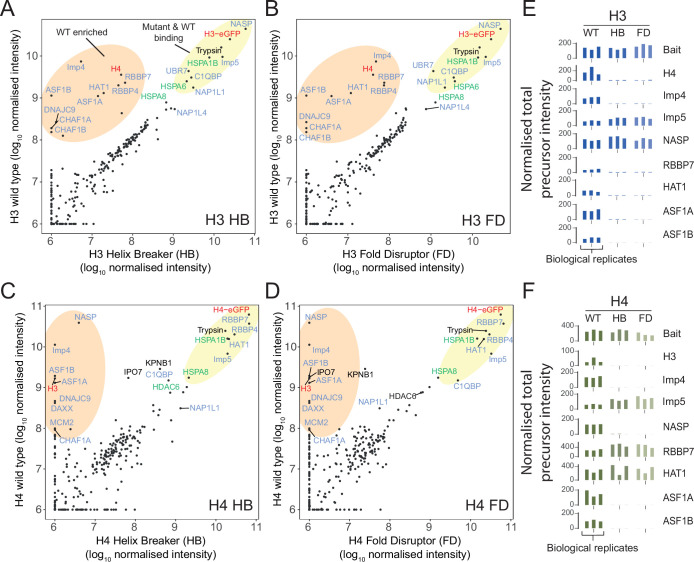
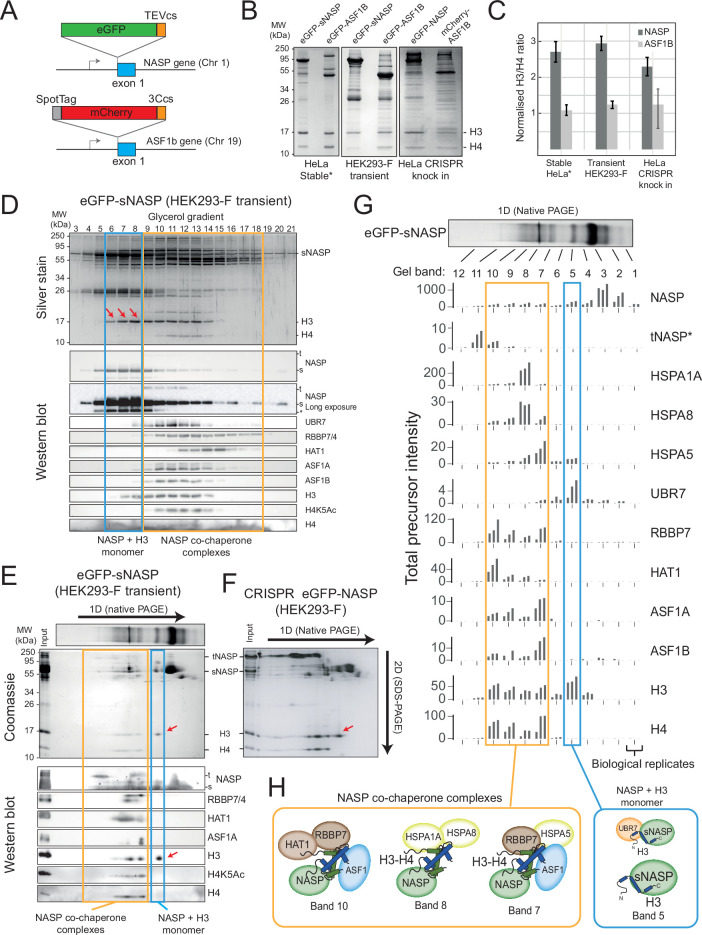
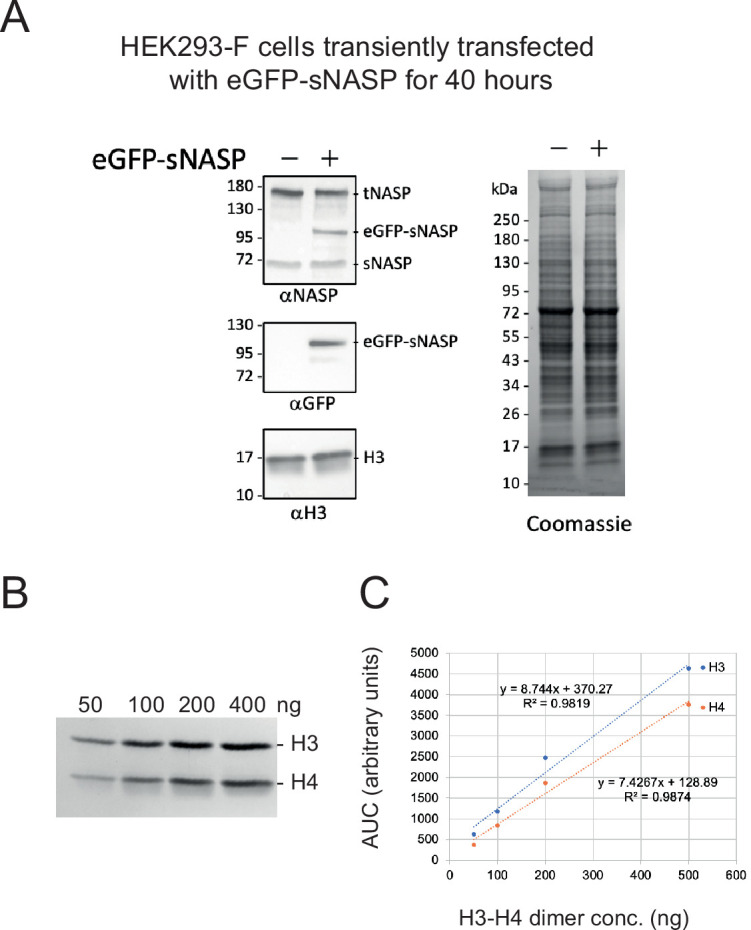
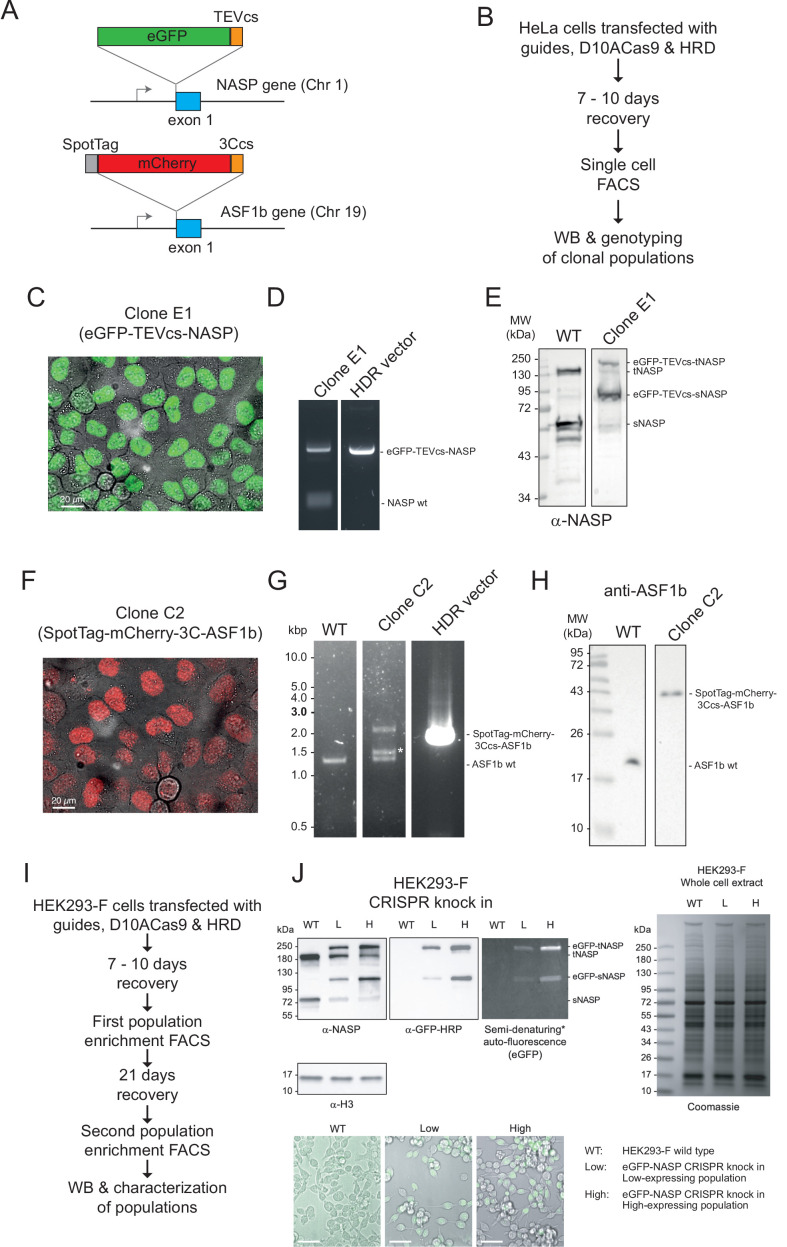
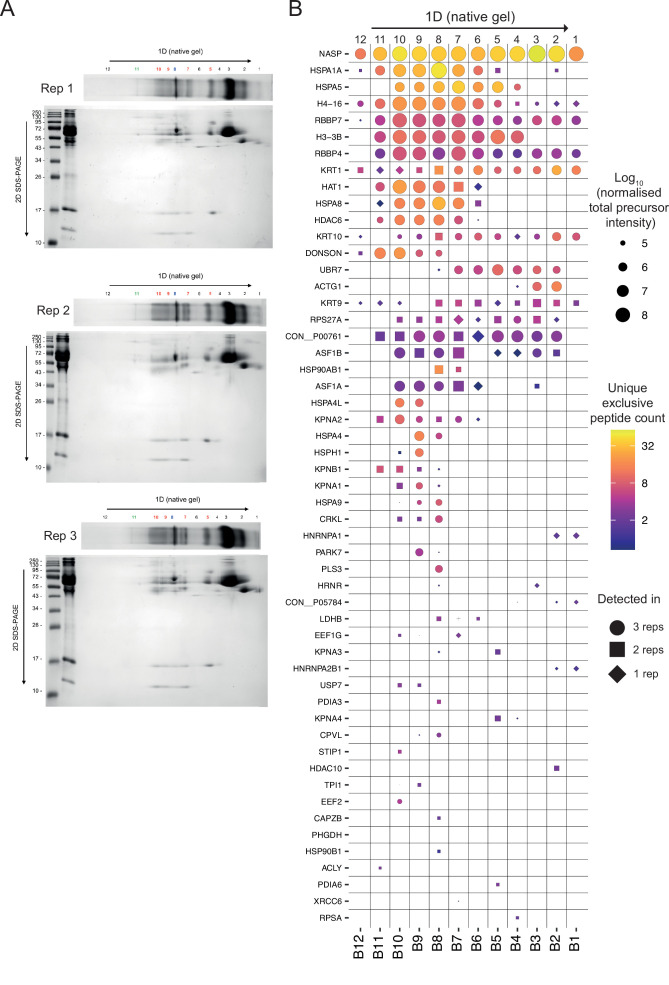
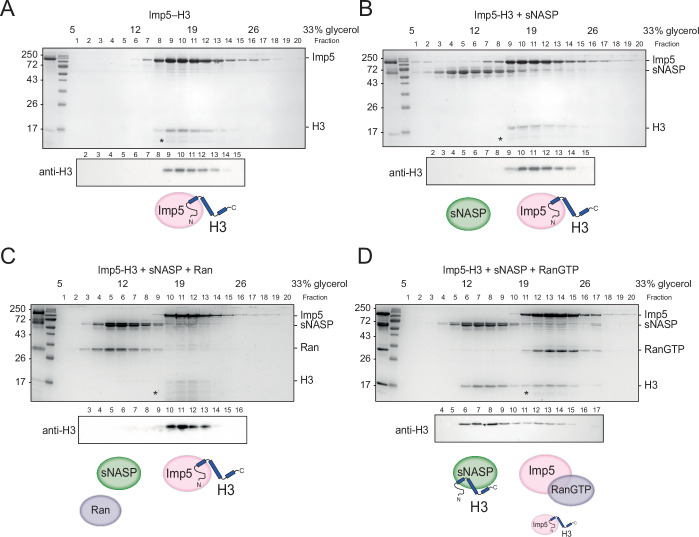
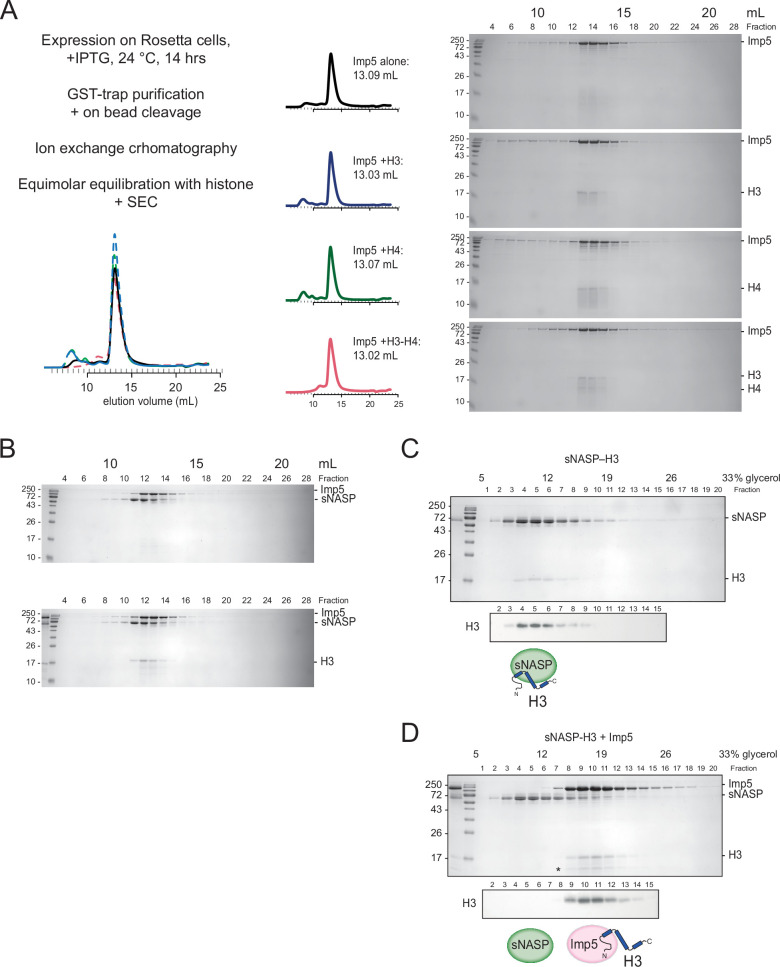
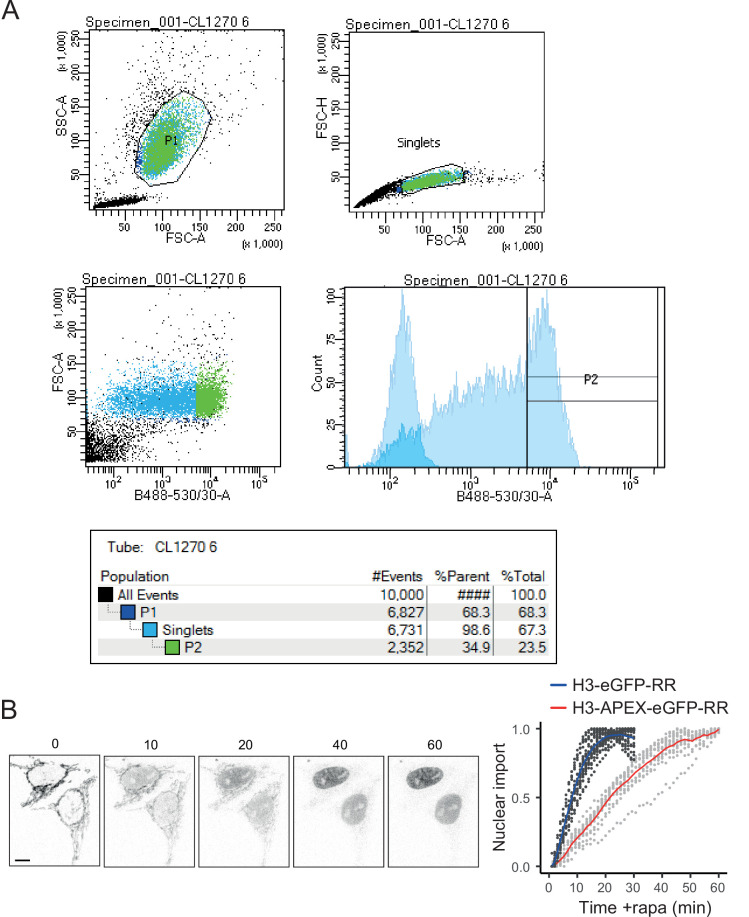
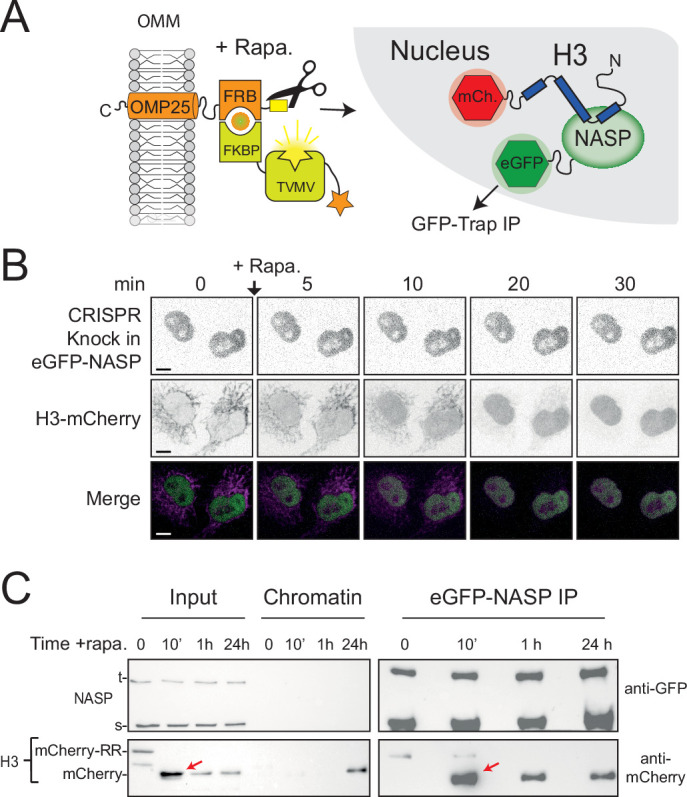
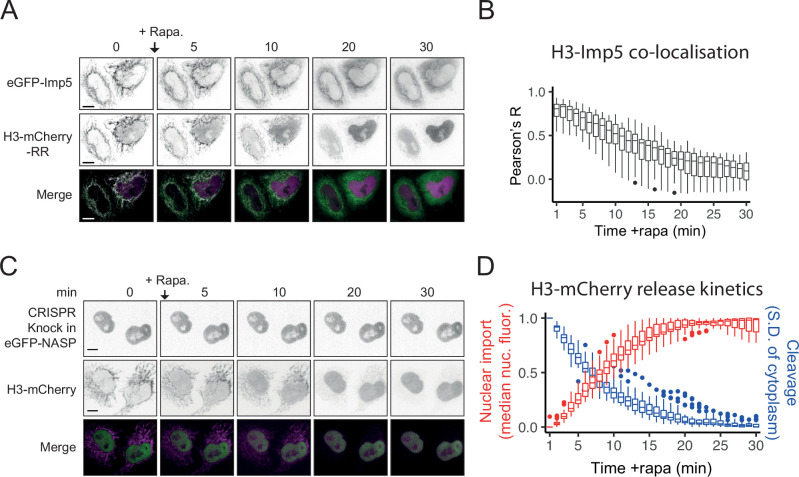
Core histones package chromosomal DNA and regulate genomic transactions, with their nuclear import and deposition involving importin-β proteins and a dedicated repertoire of histone chaperones. Previously, a histone H3-H4 dimer has been isolated bound to importin-4 (Imp4) and the chaperone ASF1, suggesting that H3 and H4 fold together in the cytoplasm before nuclear import. However, other studies have shown the existence of monomeric H3 in the nucleus, indicating a post-import folding pathway. Here, we report that the predominant importin associated with cytoplasmic H3 is importin-5 (Imp5), which hands off its monomeric cargo to nuclear sNASP. Imp5, in contrast to Imp4, binds to both H3 and H4 containing constitutively monomeric mutations and binds to newly synthesised, monomeric H3 tethered in the cytoplasm. Constitutively monomeric H3 retains its interaction with NASP, whereas monomeric H4 retains interactions specifically with HAT1 and RBBP7. High-resolution separation of NASP interactors shows the 's' isoform but not the 't' isoform associates with monomeric H3, whilst both isoforms associate with H3-H4 dimers in at least three discrete multi-chaperoning complexes. In vitro binding experiments show mutual exclusivity between sNASP and Imp5 in binding H3, suggesting direct competition for interaction sites, with the GTP-bound form of Ran required for histone transfer. Finally, using pulse-chase analysis, we show that cytoplasm-tethered histones do not interact with endogenous NASP until they reach the nucleus, whereupon they bind rapidly. We propose an Imp5-specific import pathway for monomeric H3 that hands off to sNASP in the nucleus, with a parallel H4 pathway involving Imp5 and the HAT1-RBBP7 complex, followed by nuclear folding and hand-off to deposition factors.
核心组蛋白包裹染色体 DNA 并调节基因组的各种活动,其核输入和沉积涉及输入蛋白-β 和一组专门的组蛋白伴侣。以前,已分离出与输入蛋白 4(Imp4)和伴侣 ASF1 结合的组蛋白 H3-H4 二聚体,表明 H3 和 H4 在核输入之前在细胞质中折叠在一起。然而,其他研究表明核内存在单体 H3,表明存在核输入后的折叠途径。在这里,我们报告说与细胞质 H3 相关的主要输入蛋白是输入蛋白 5(Imp5),它将其单体货物传递给核 sNASP。与 Imp4 相反,Imp5 与含有组成型单体突变的 H3 和 H4 结合,并与细胞质中被束缚的新合成的单体 H3 结合。组成型单体 H3 保持与 NASP 的相互作用,而单体 H4 保留与 HAT1 和 RBBP7 的特异性相互作用。NASP 相互作用物的高分辨率分离显示出“s”同工型而不是“t”同工型与单体 H3 结合,而两种同工型至少在三个离散的多伴侣复合物中与 H3-H4 二聚体结合。体外结合实验表明 sNASP 和 Imp5 在结合 H3 时具有互斥性,表明直接竞争相互作用位点,Histone transfer 需要 Ran 的 GTP 结合形式。最后,通过脉冲追踪分析,我们表明细胞质束缚的组蛋白在到达细胞核之前不会与内源性 NASP 相互作用,一旦进入细胞核,它们就会迅速结合。我们提出了一种 Imp5 特异性的单体 H3 输入途径,该途径在细胞核中传递给 sNASP,同时涉及 Imp5 和 HAT1-RBBP7 复合物的平行 H4 途径,随后进行核折叠并传递给沉积因子。