Tri-Institutional PhD Program in Computational Biology and Medicine, Weill Cornell Medicine, New York, NY, USA.
Computational Oncology, Department of Epidemiology and Biostatistics, Memorial Sloan Kettering Cancer Center, New York, NY, USA.
Nature. 2022 Dec;612(7938):106-115. doi: 10.1038/s41586-022-05249-0. Epub 2022 Oct 26.
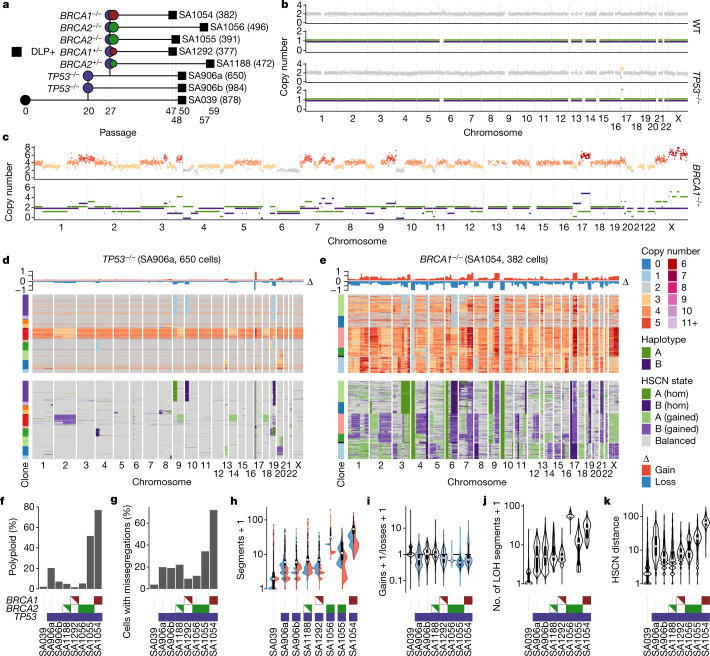
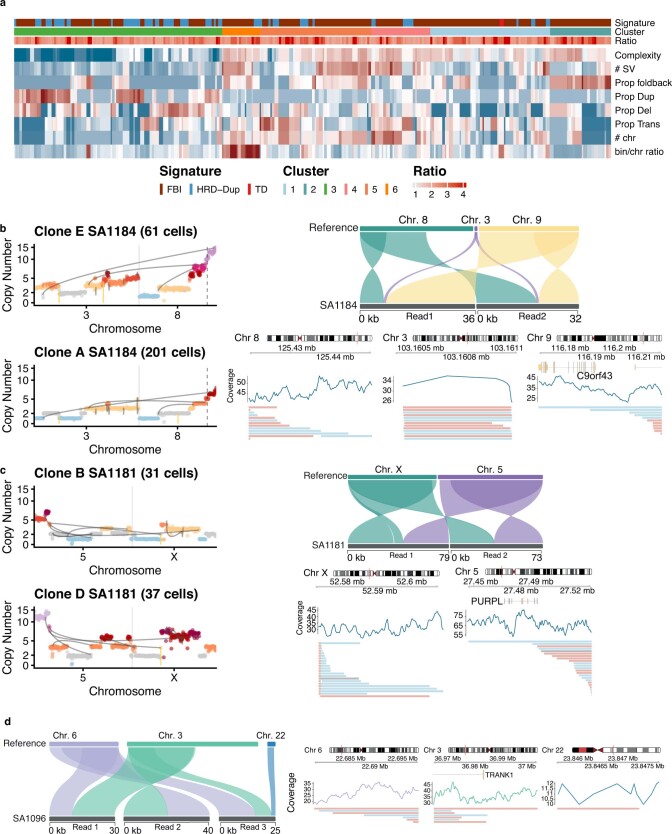
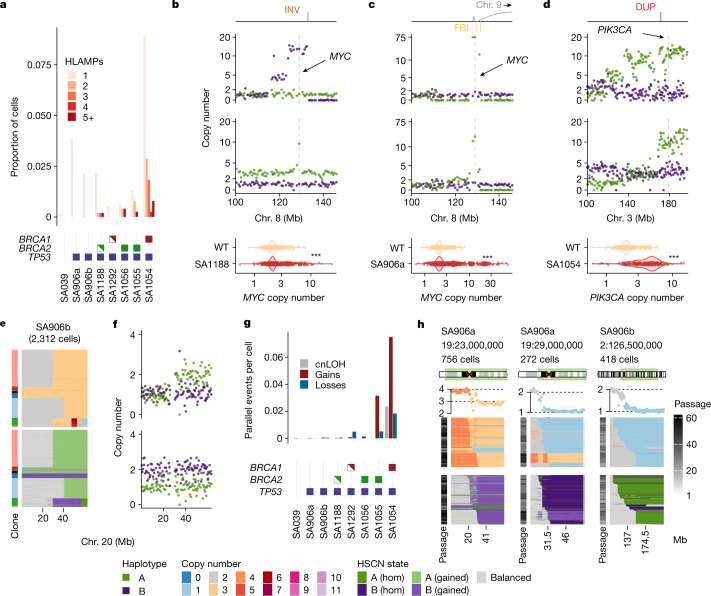
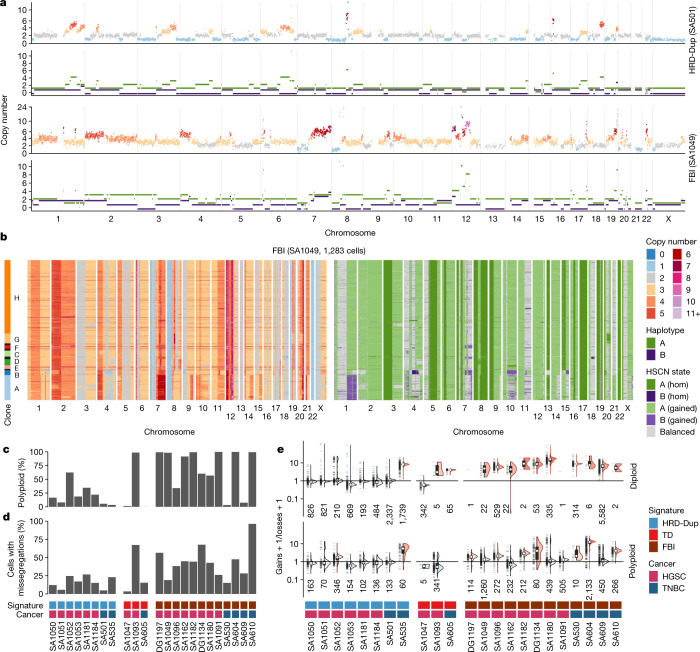
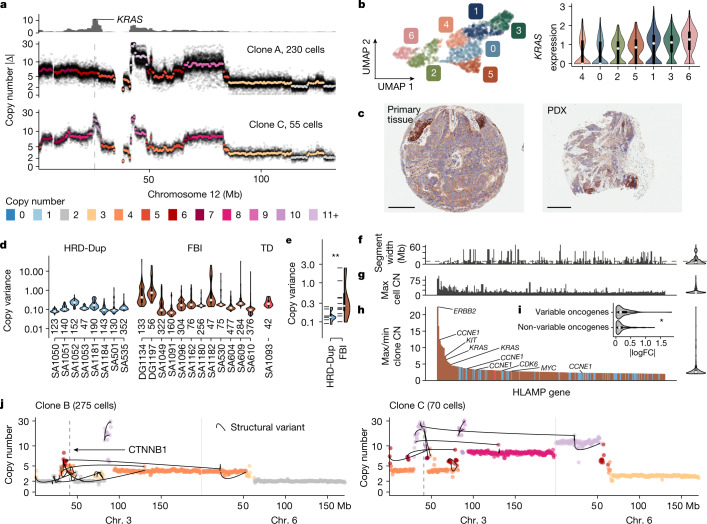
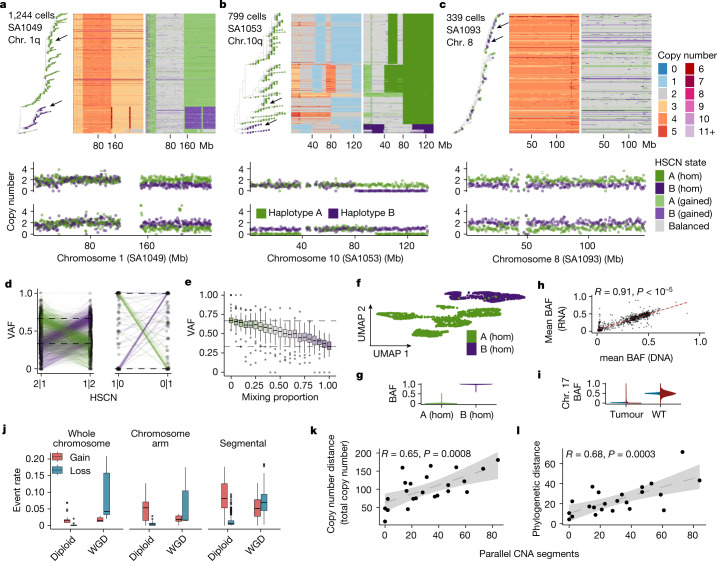
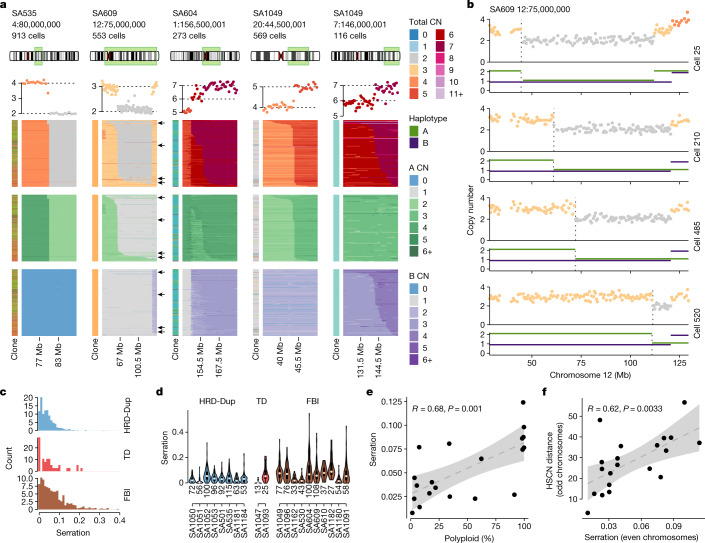
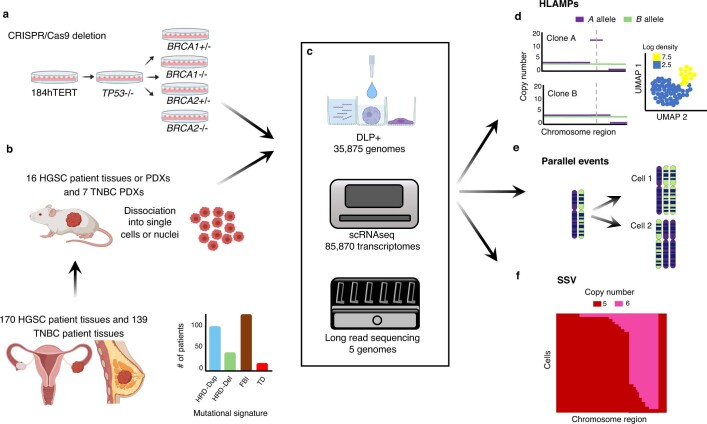
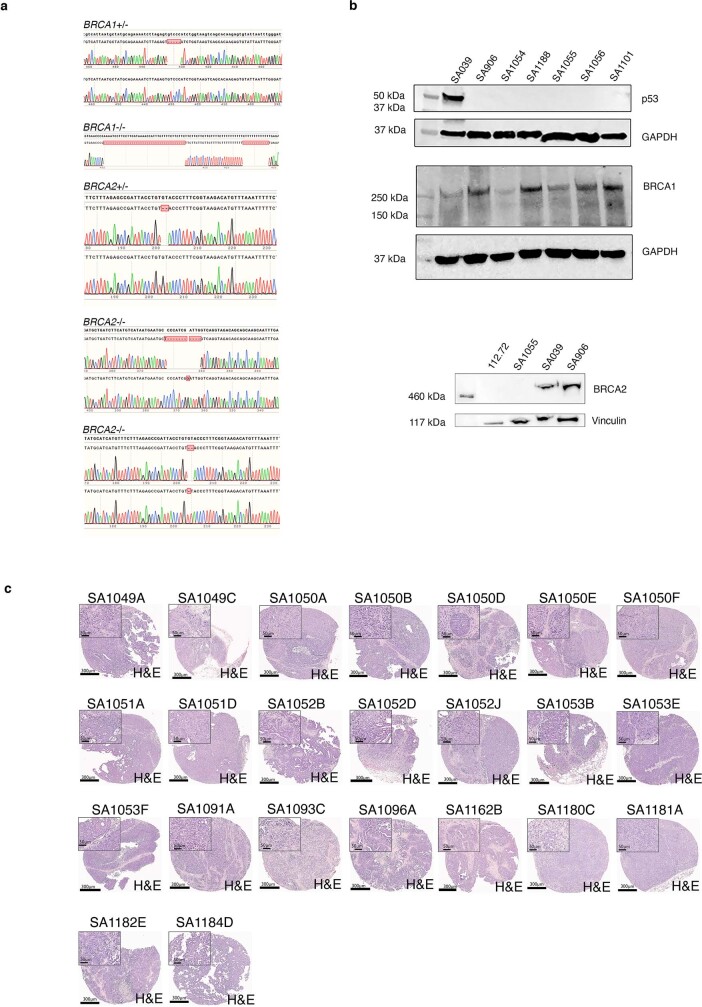
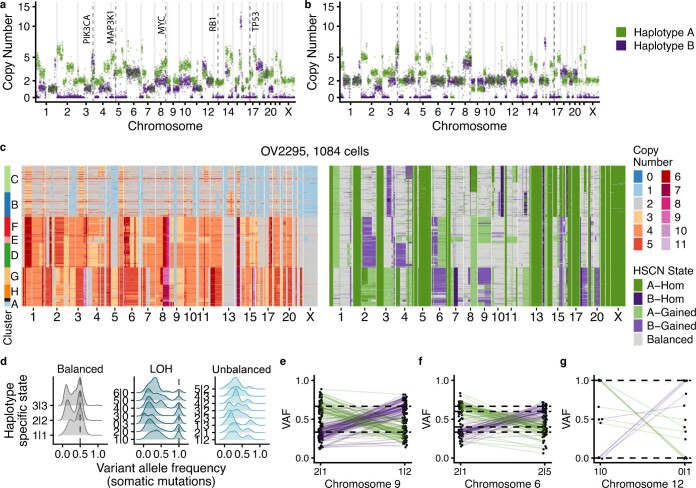
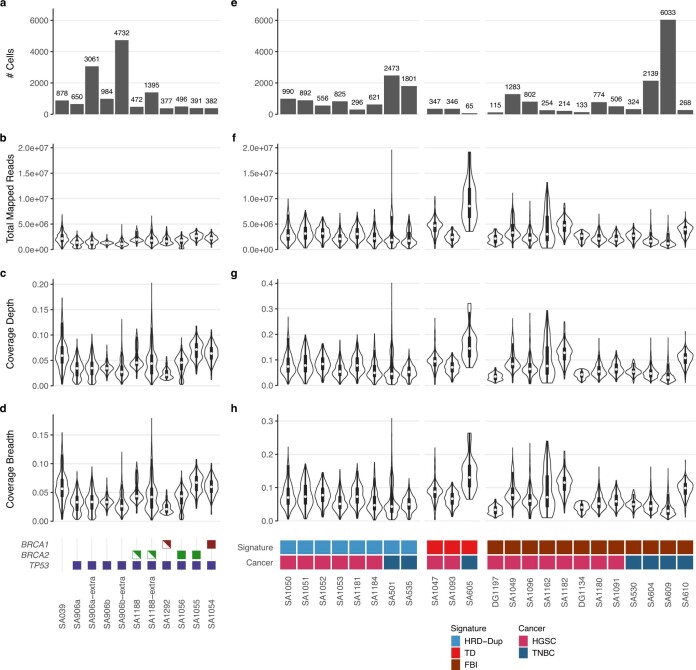
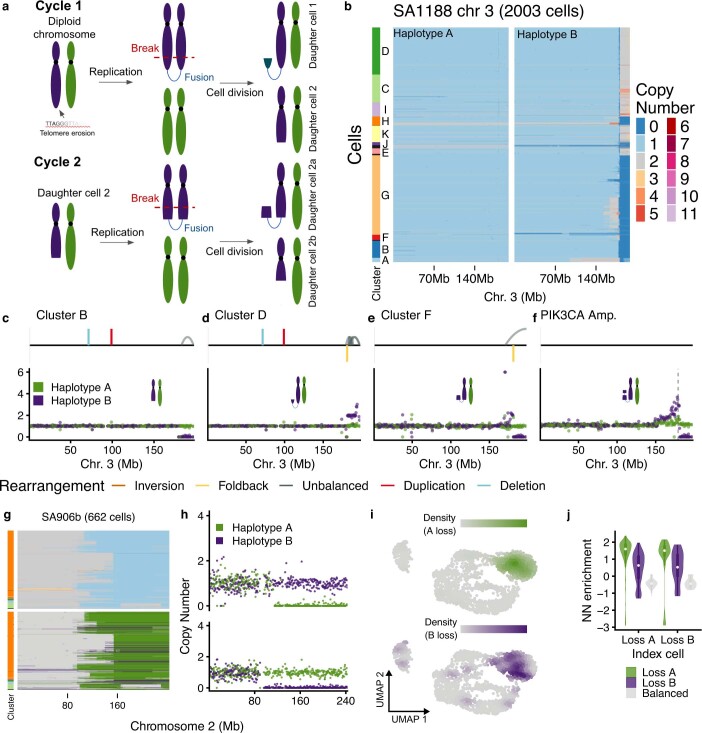
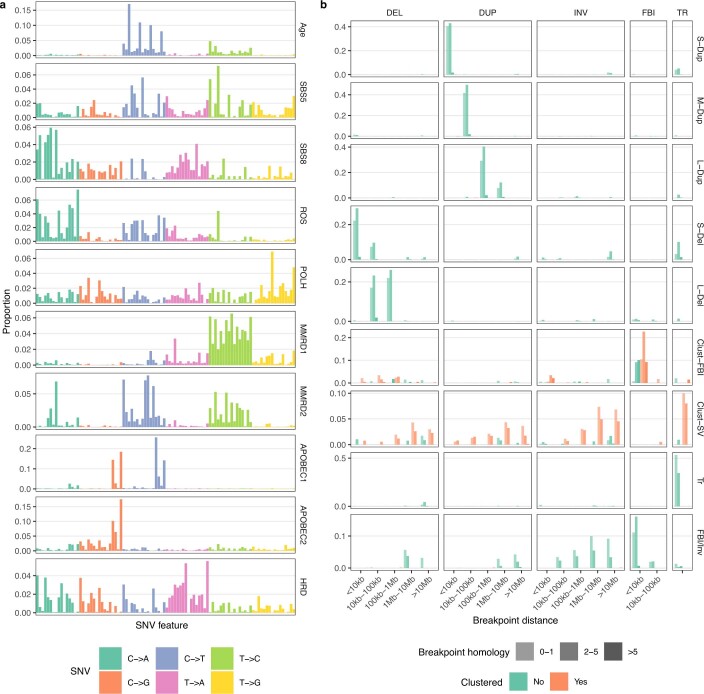
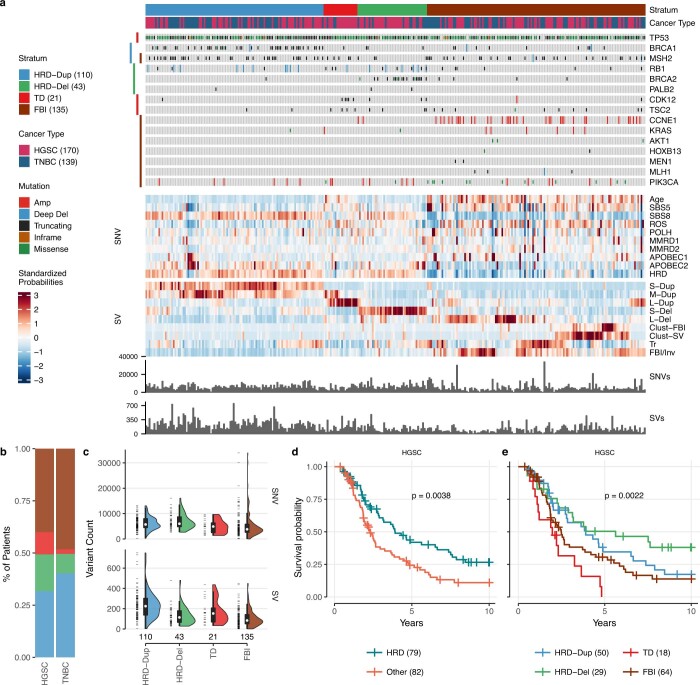
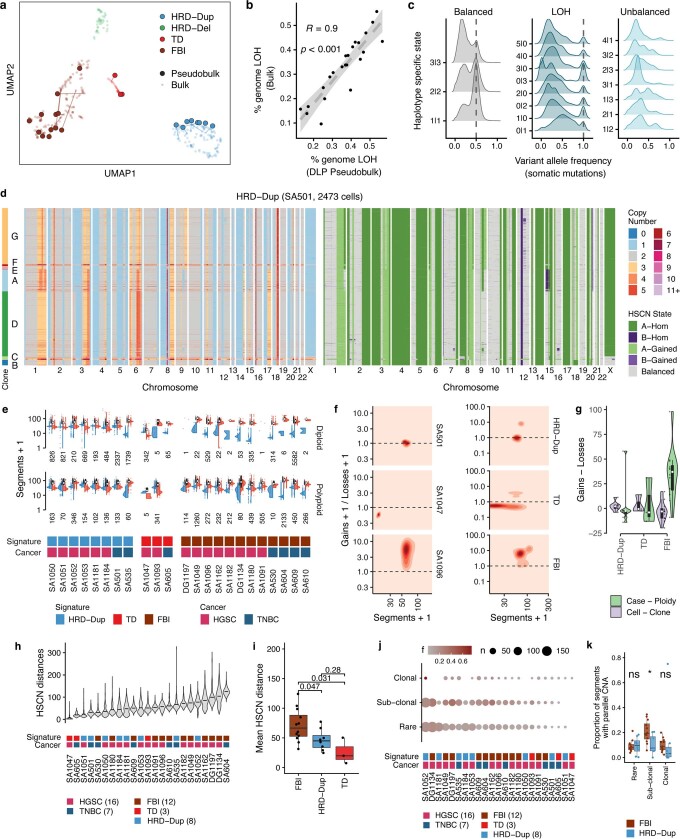
How cell-to-cell copy number alterations that underpin genomic instability in human cancers drive genomic and phenotypic variation, and consequently the evolution of cancer, remains understudied. Here, by applying scaled single-cell whole-genome sequencing to wild-type, TP53-deficient and TP53-deficient;BRCA1-deficient or TP53-deficient;BRCA2-deficient mammary epithelial cells (13,818 genomes), and to primary triple-negative breast cancer (TNBC) and high-grade serous ovarian cancer (HGSC) cells (22,057 genomes), we identify three distinct 'foreground' mutational patterns that are defined by cell-to-cell structural variation. Cell- and clone-specific high-level amplifications, parallel haplotype-specific copy number alterations and copy number segment length variation (serrate structural variations) had measurable phenotypic and evolutionary consequences. In TNBC and HGSC, clone-specific high-level amplifications in known oncogenes were highly prevalent in tumours bearing fold-back inversions, relative to tumours with homologous recombination deficiency, and were associated with increased clone-to-clone phenotypic variation. Parallel haplotype-specific alterations were also commonly observed, leading to phylogenetic evolutionary diversity and clone-specific mono-allelic expression. Serrate variants were increased in tumours with fold-back inversions and were highly correlated with increased genomic diversity of cellular populations. Together, our findings show that cell-to-cell structural variation contributes to the origins of phenotypic and evolutionary diversity in TNBC and HGSC, and provide insight into the genomic and mutational states of individual cancer cells.
细胞间拷贝数改变如何导致人类癌症中的基因组不稳定性,进而驱动基因组和表型的变异,并推动癌症的进化,这方面的研究仍不够充分。在这里,我们通过对野生型、TP53 缺陷型和 TP53 缺陷型;BRCA1 缺陷型或 TP53 缺陷型;BRCA2 缺陷型乳腺上皮细胞(13818 个基因组),以及原发性三阴性乳腺癌(TNBC)和高级别浆液性卵巢癌(HGSC)细胞(22057 个基因组)进行规模化单细胞全基因组测序,鉴定出三种不同的“突出”突变模式,这些模式由细胞间结构变异定义。细胞和克隆特异性高水平扩增、平行单倍型特异性拷贝数改变和拷贝数片段长度变异(锯齿状结构变异)具有可测量的表型和进化后果。在 TNBC 和 HGSC 中,已知致癌基因的克隆特异性高水平扩增在具有折返倒位的肿瘤中比具有同源重组缺陷的肿瘤更为常见,并且与克隆间表型变异增加相关。平行单倍型特异性改变也经常观察到,导致系统发育进化多样性和克隆特异性单等位基因表达。锯齿状变异在具有折返倒位的肿瘤中增加,并且与细胞群体的基因组多样性增加高度相关。总之,我们的研究结果表明,细胞间结构变异有助于 TNBC 和 HGSC 中表型和进化多样性的起源,并为了解单个癌细胞的基因组和突变状态提供了线索。