Department of Biology and Center for Genomics and Systems Biology, New York University, New York, United States.
UCSF HSW, San Francisco, United States.
Elife. 2022 Nov 4;11:e68745. doi: 10.7554/eLife.68745.
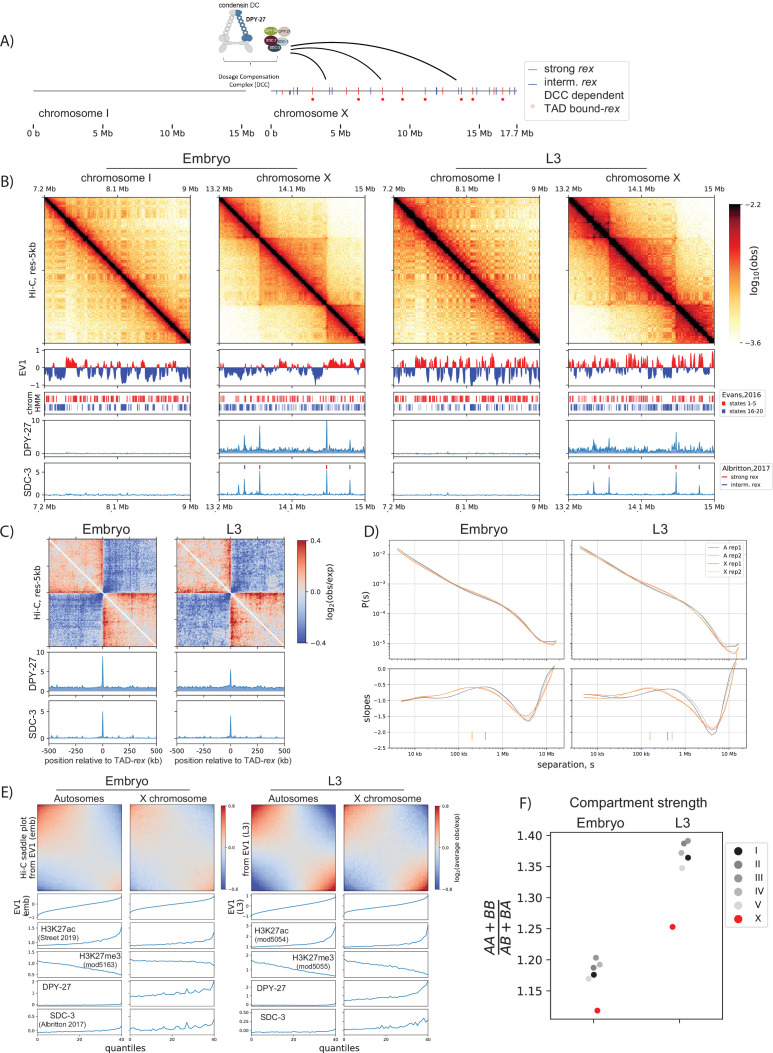
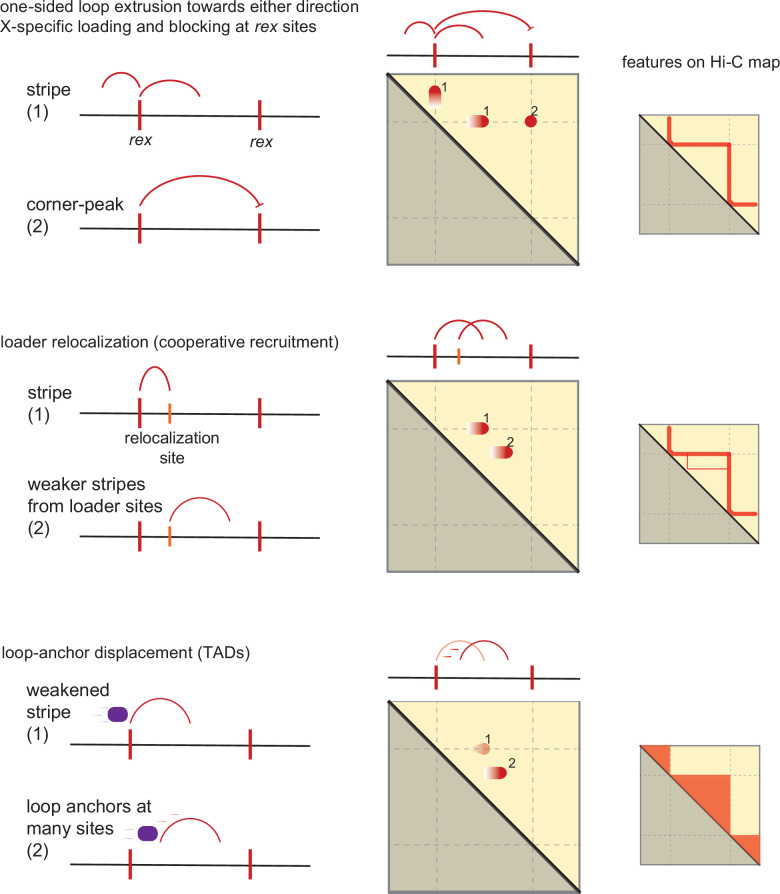
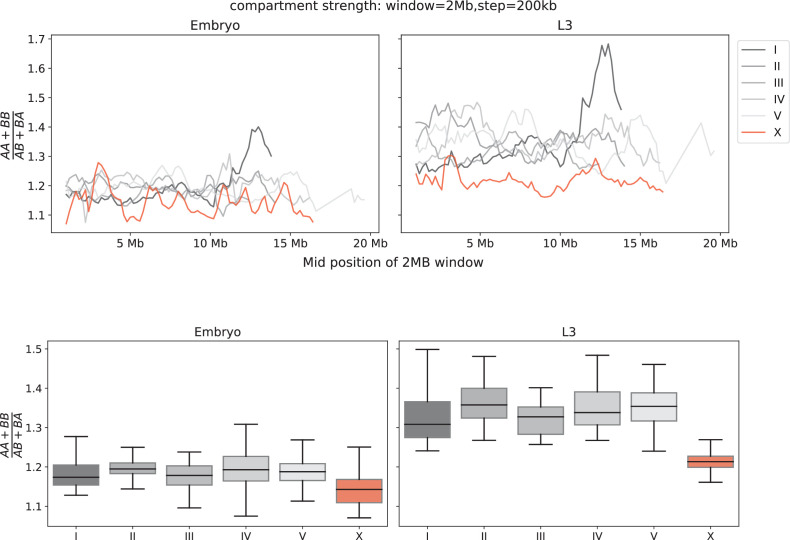
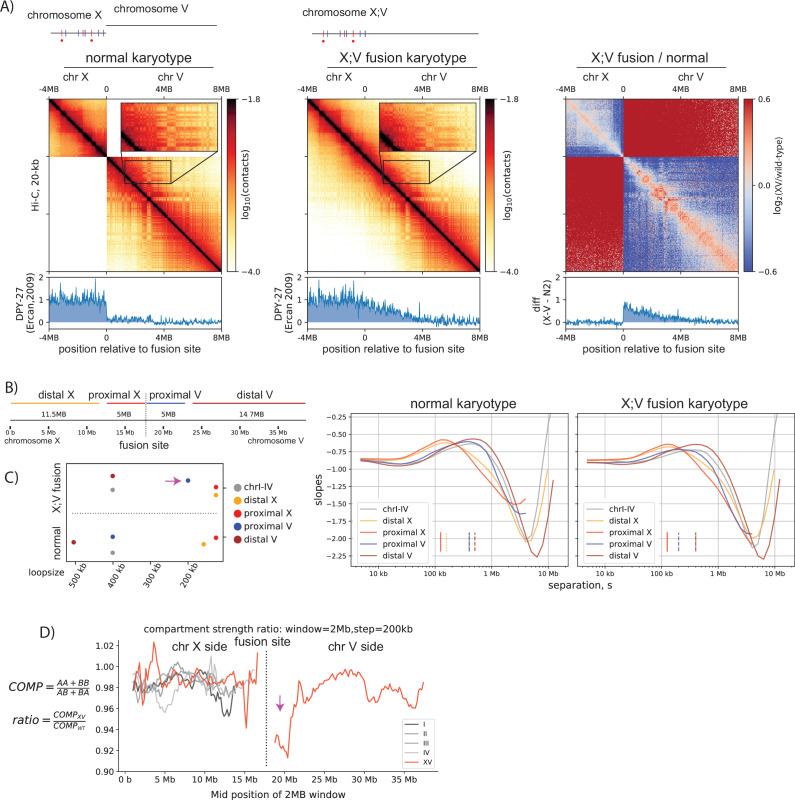
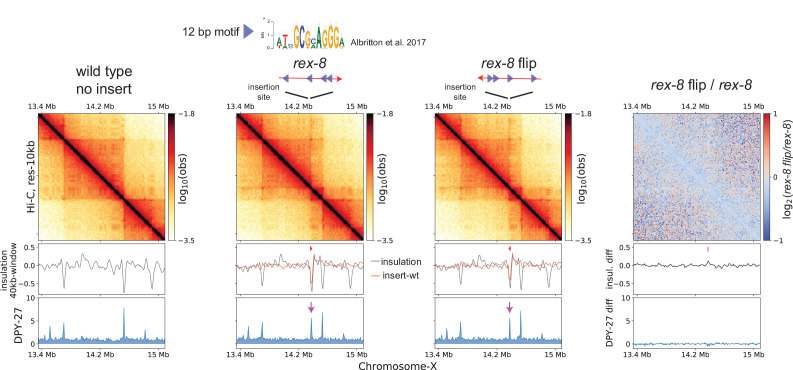
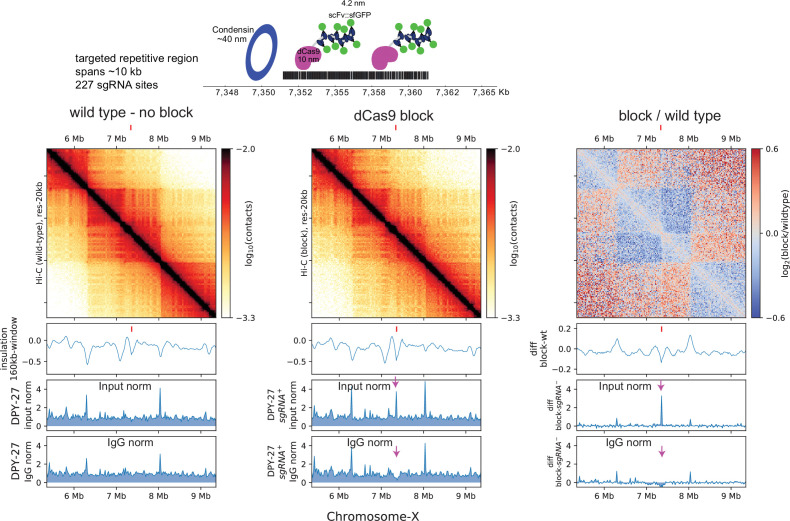
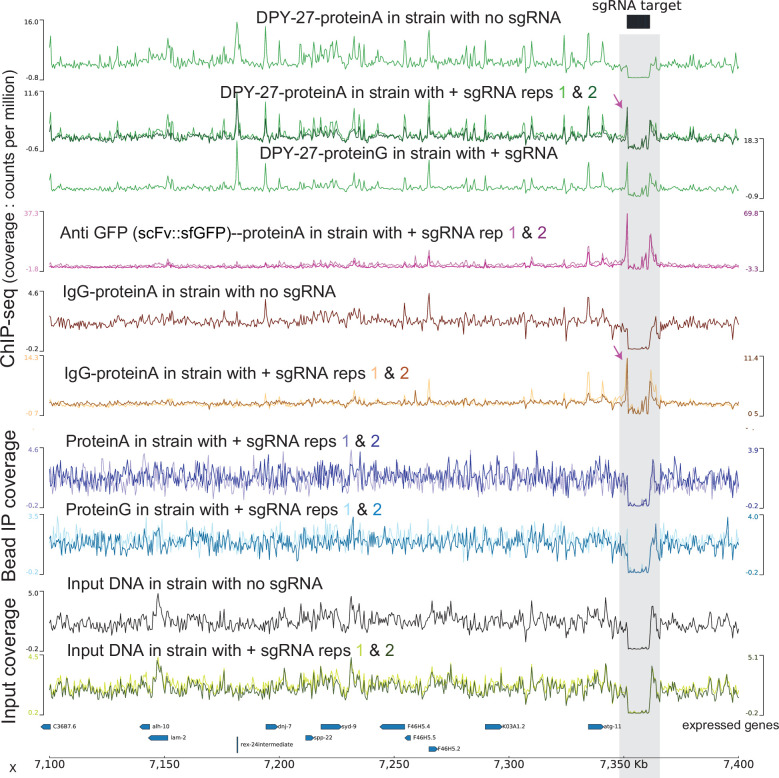
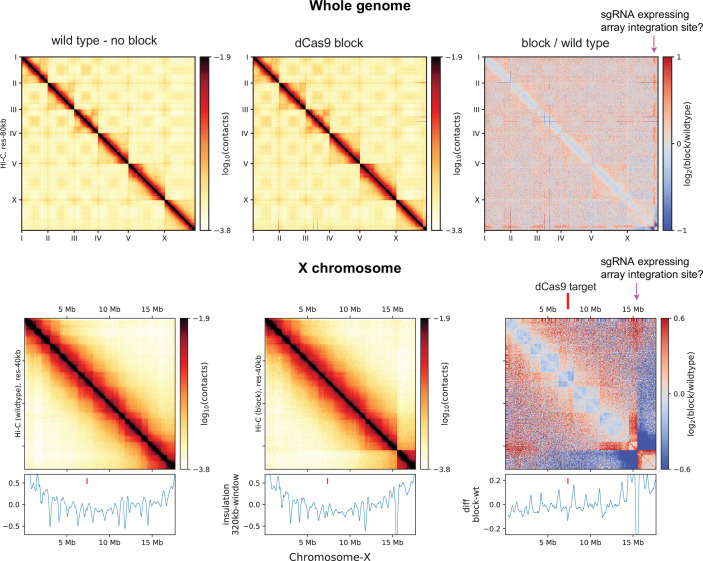
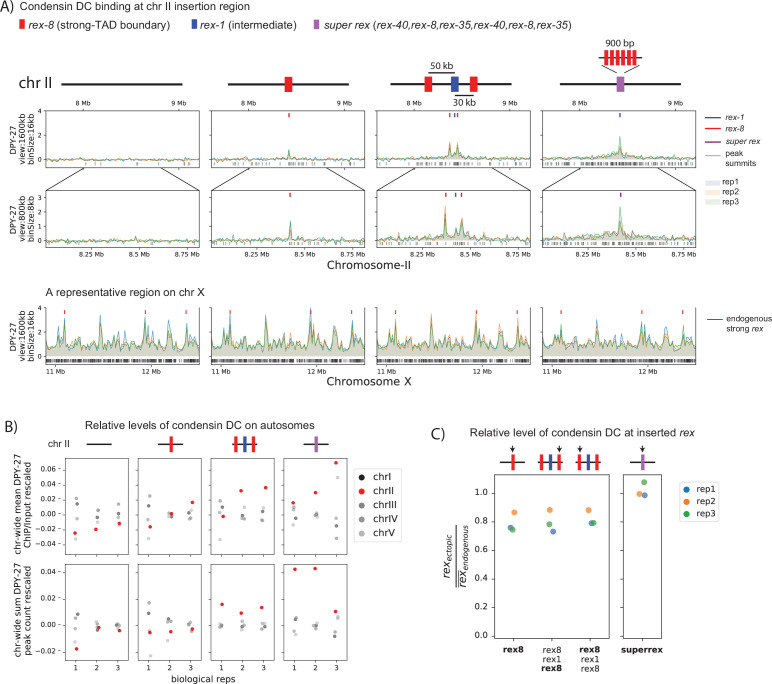
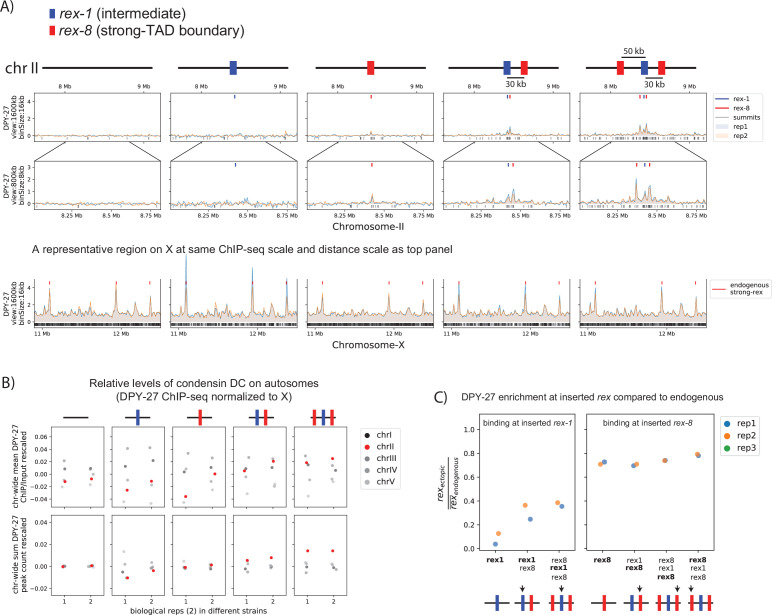
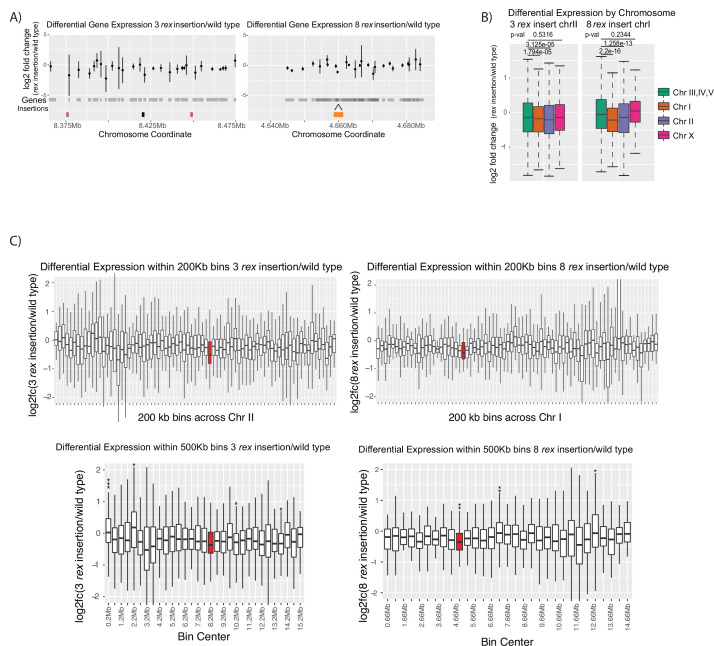
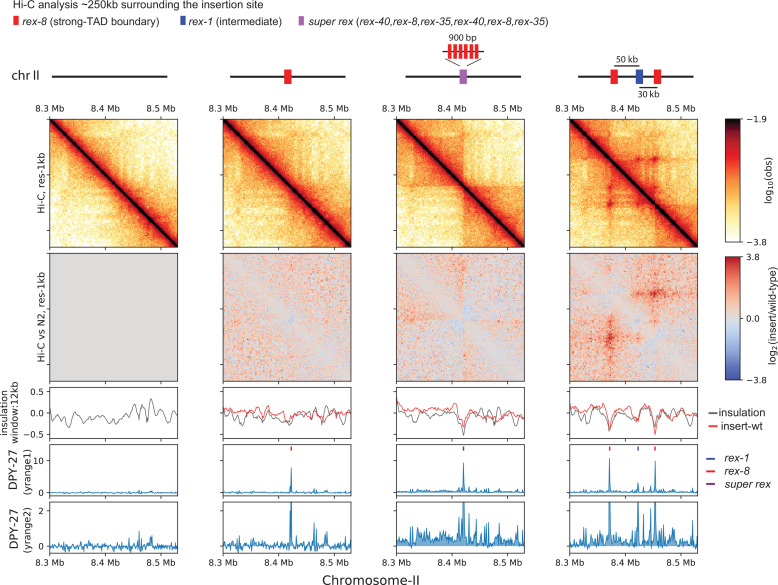
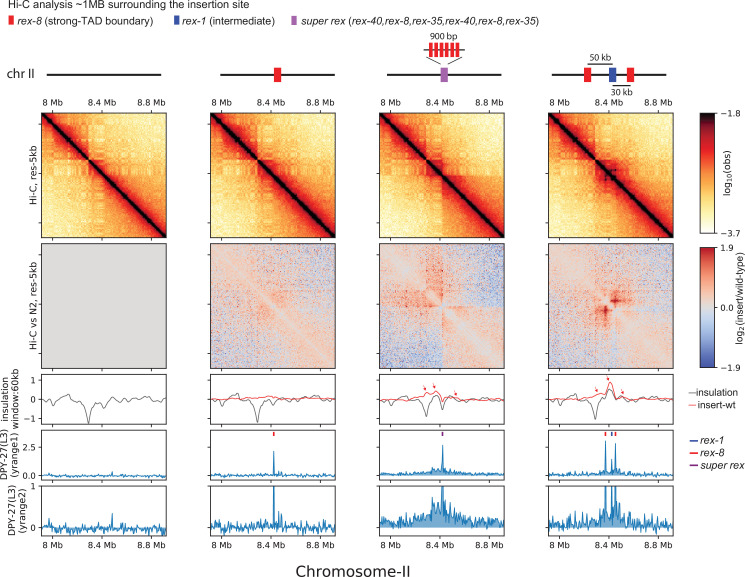
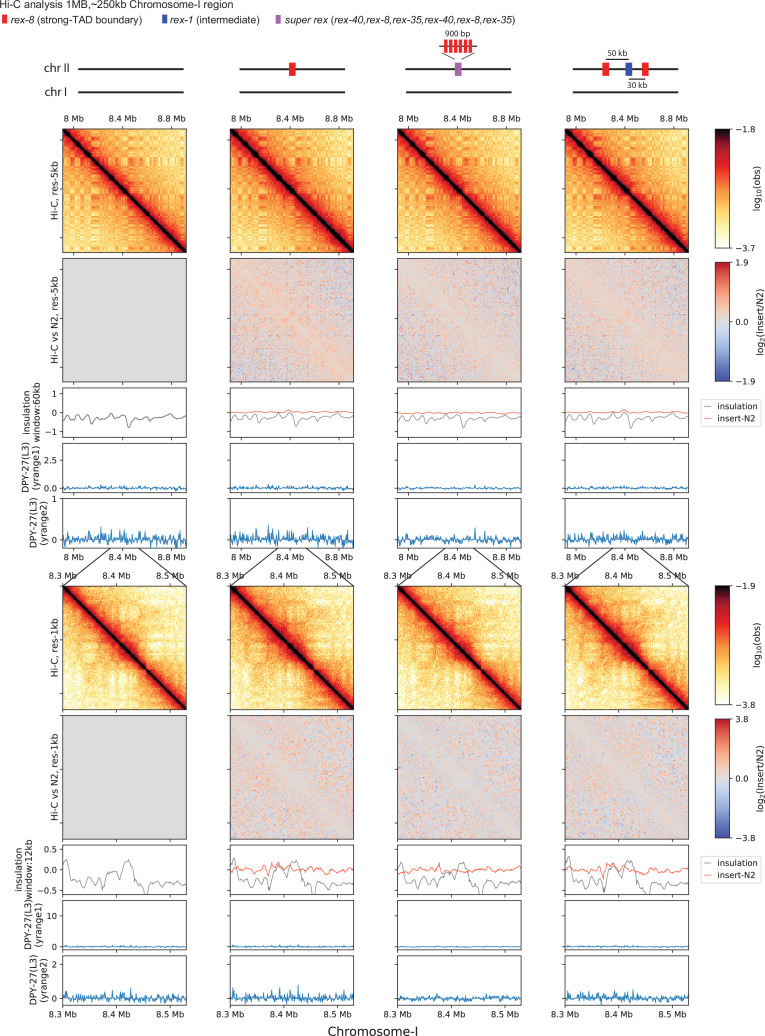
Condensins are molecular motors that compact DNA via linear translocation. In , the X-chromosome harbors a specialized condensin that participates in dosage compensation (DC). Condensin DC is recruited to and spreads from a small number of ecruitment lements on the -chromosome () and is required for the formation of topologically associating domains (TADs). We take advantage of autosomes that are largely devoid of condensin DC and TADs to address how sites and condensin DC give rise to the formation of TADs. When an autosome and X-chromosome are physically fused, despite the spreading of condensin DC into the autosome, no TAD was created. Insertion of a strong on the X-chromosome results in the TAD boundary formation regardless of sequence orientation. When the same is inserted on an autosome, despite condensin DC recruitment, there was no spreading or features of a TAD. On the other hand, when a '' composed of six sites or three separate sites are inserted on an autosome, recruitment and spreading of condensin DC led to the formation of TADs. Therefore, recruitment to and spreading from sites are necessary and sufficient for recapitulating loop-anchored TADs observed on the X-chromosome. Together our data suggest a model in which sites are both loading sites and bidirectional barriers for condensin DC, a one-sided loop-extruder with movable inactive anchor.
凝缩蛋白是通过线性易位来压缩 DNA 的分子马达。在 中,X 染色体上有一种特殊的凝缩蛋白,参与剂量补偿(DC)。凝缩蛋白 DC 被招募到并从 -染色体上的少数招募元件( )扩散,并需要形成拓扑关联域(TAD)。我们利用基本上缺乏凝缩蛋白 DC 和 TAD 的常染色体来解决 位点和凝缩蛋白 DC 如何导致 TAD 形成的问题。当一条常染色体和 X 染色体物理融合时,尽管凝缩蛋白 DC 扩散到常染色体中,但没有形成 TAD。在 X 染色体上插入一个强 ,无论序列取向如何,都会导致 TAD 边界的形成。当相同的 插入常染色体时,尽管招募了凝缩蛋白 DC,但没有扩散或 TAD 的特征。另一方面,当六个 位点或三个独立的 位点组成的“ ”插入常染色体时,凝缩蛋白 DC 的招募和扩散导致了 TAD 的形成。因此,招募到并从 位点扩散是重现已在 X 染色体上观察到的环锚定 TAD 所必需和充分的条件。我们的数据共同表明, 位点既是凝缩蛋白 DC 的加载位点,也是双向障碍,是一种具有可移动非活性锚的单侧环挤出器。