Department of Molecular Biology, Princeton University, Princeton, United States.
Elife. 2023 Jan 4;12:e78188. doi: 10.7554/eLife.78188.
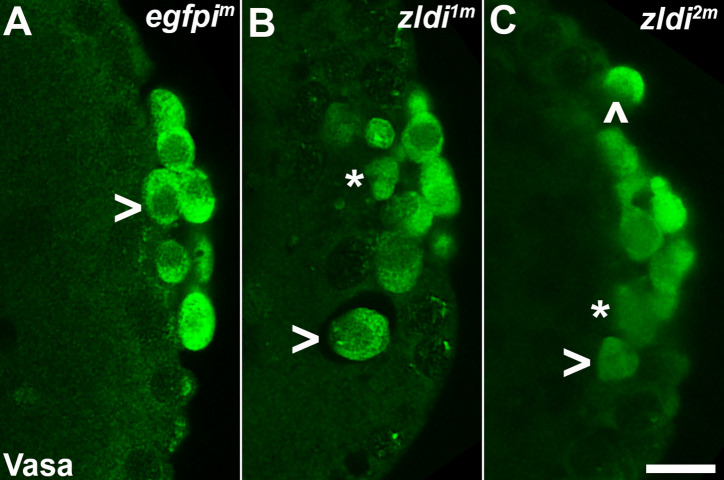
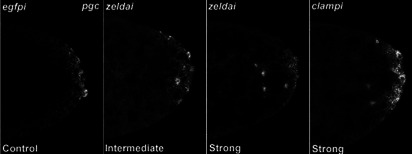
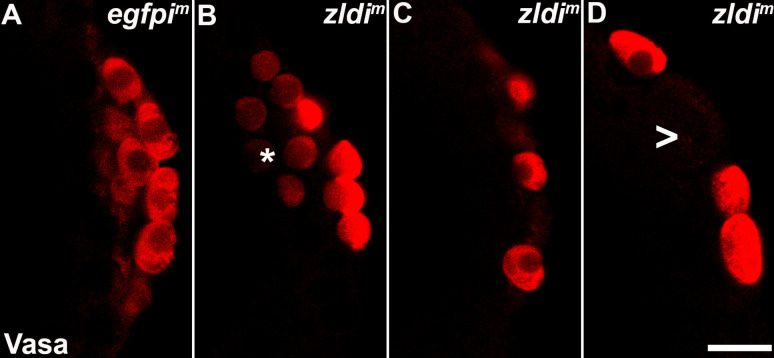
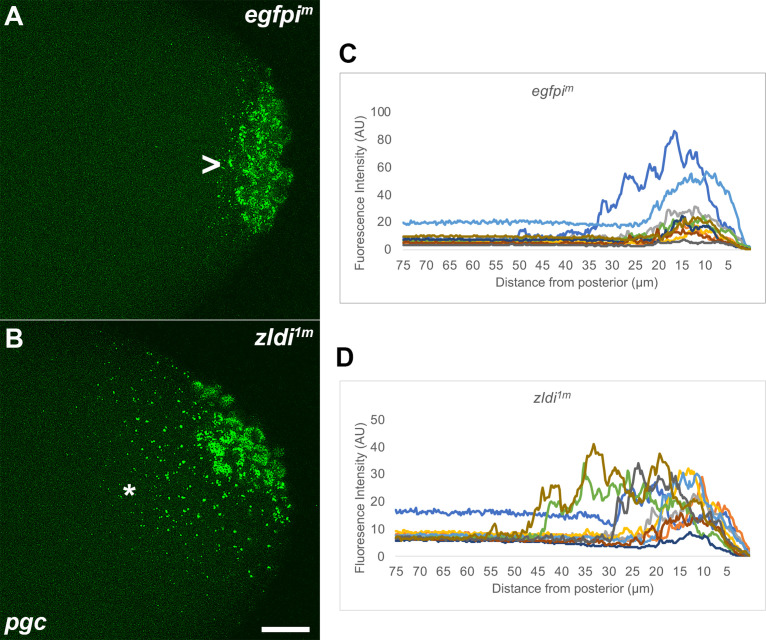
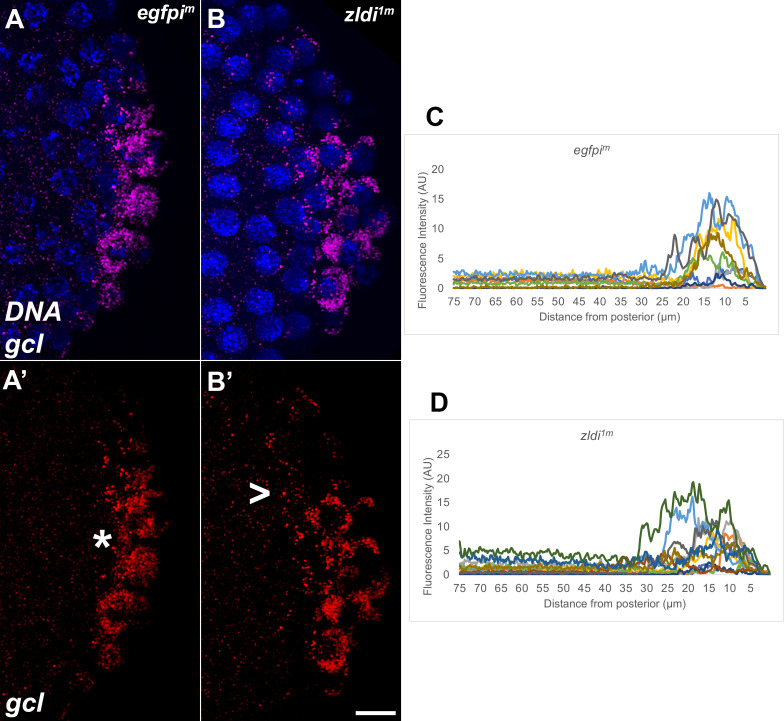
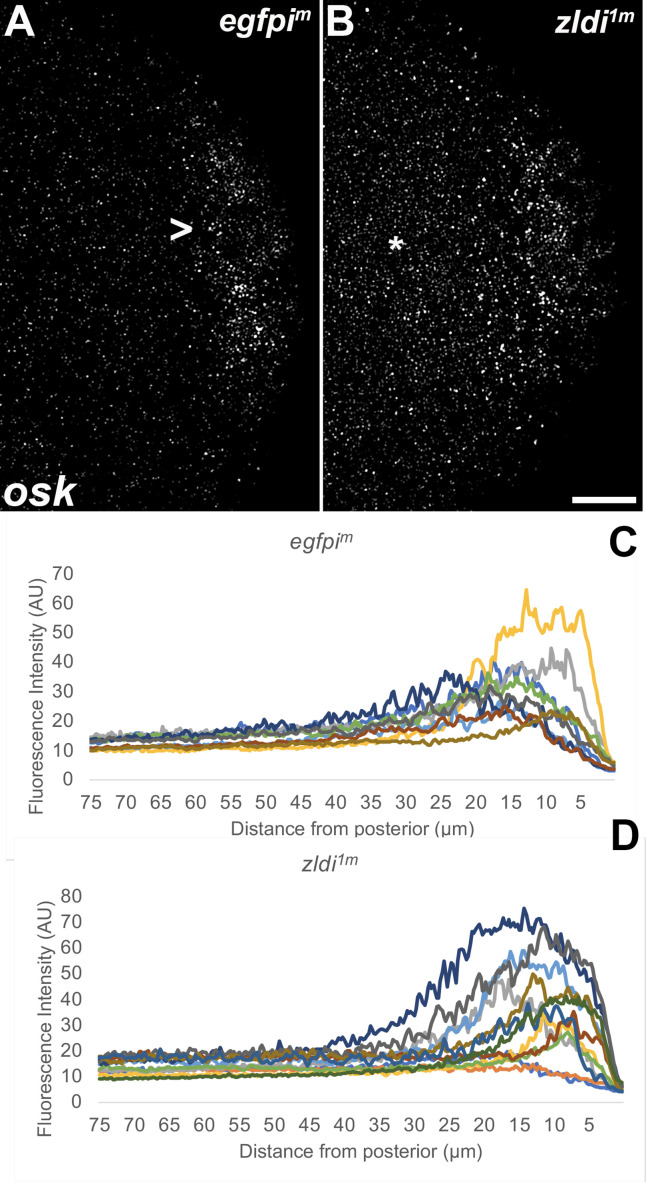
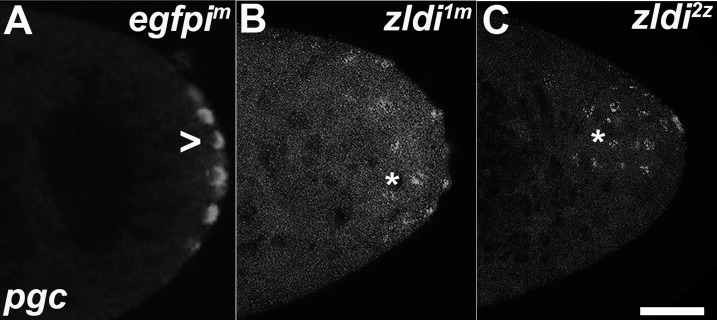
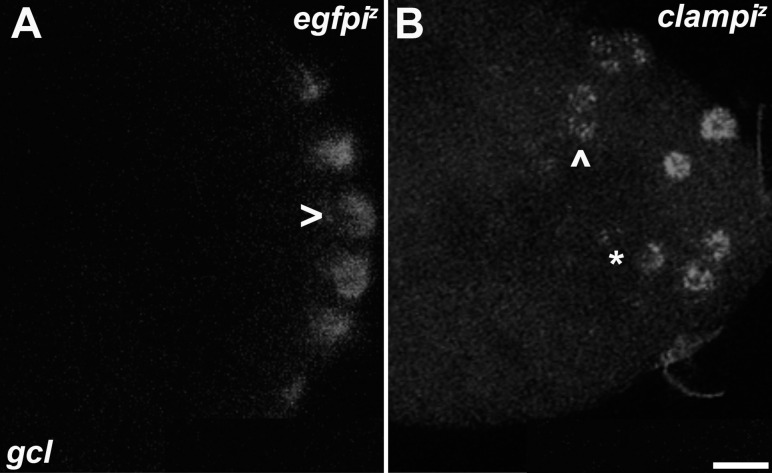
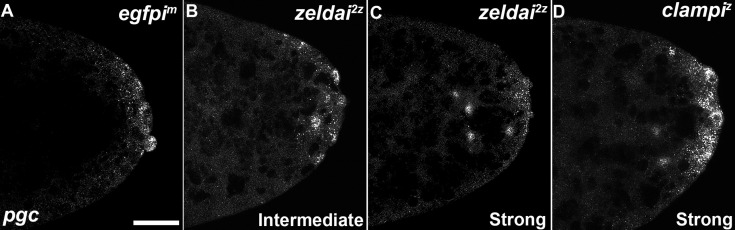
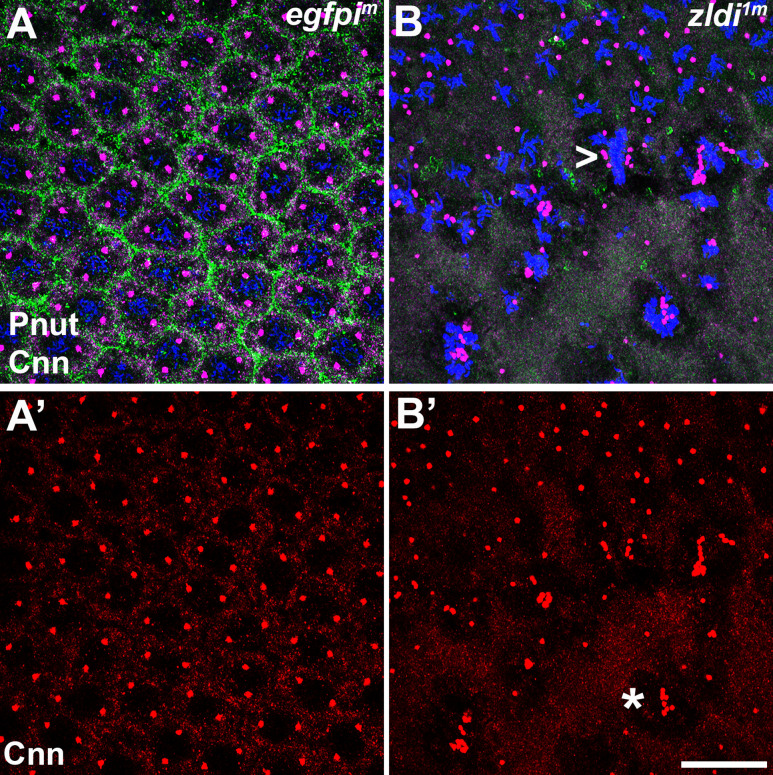
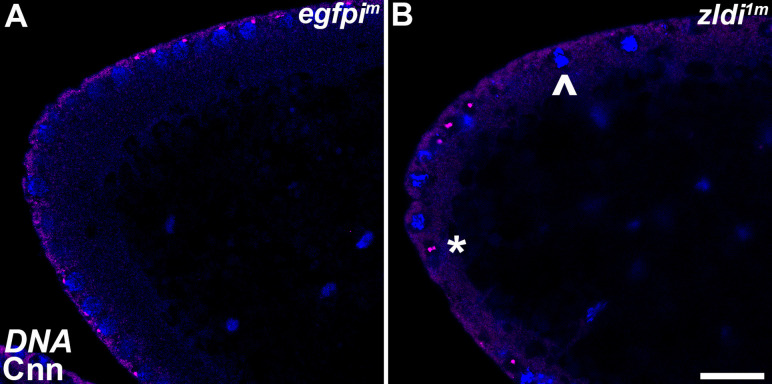
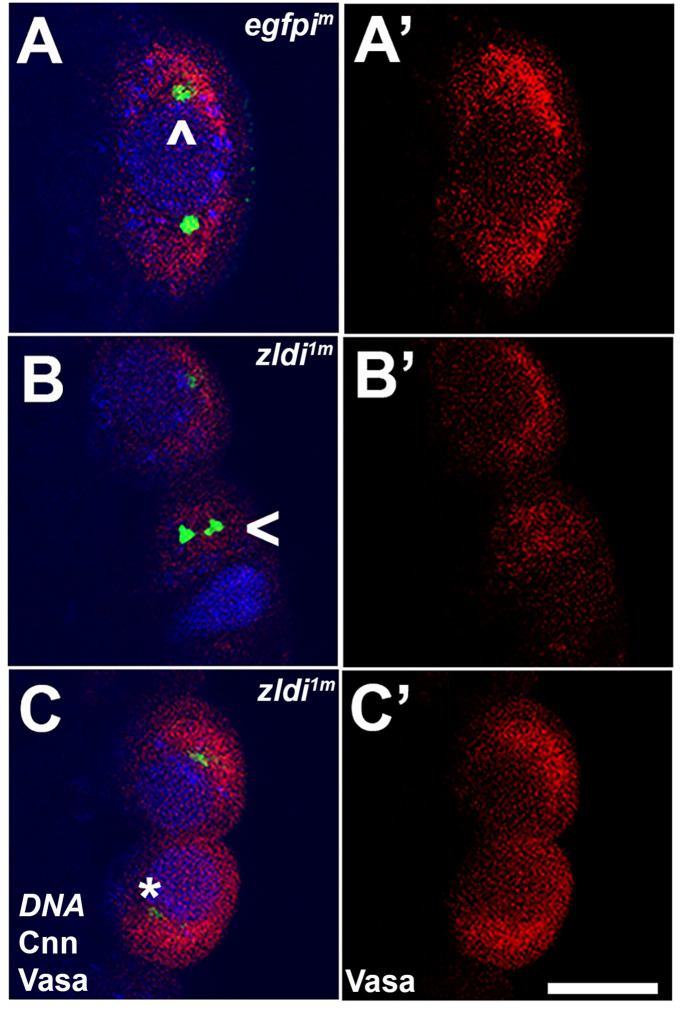
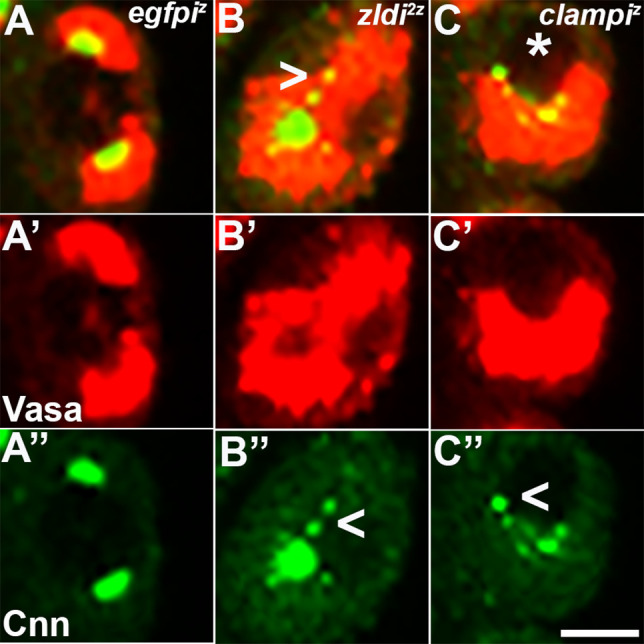
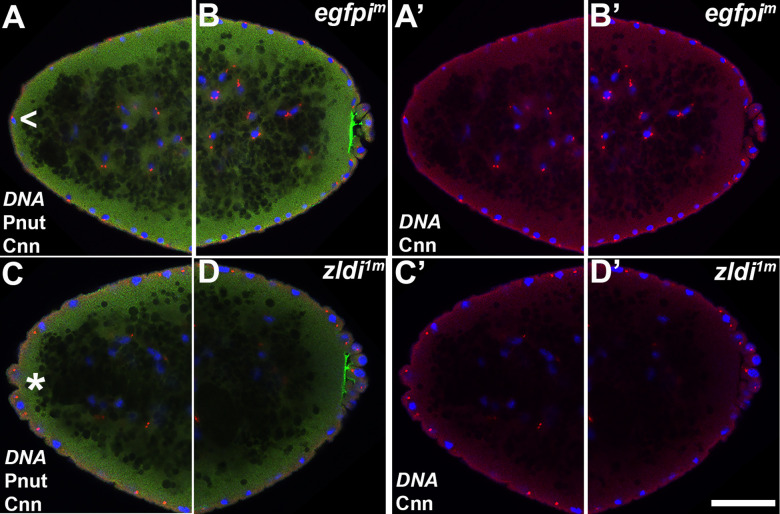
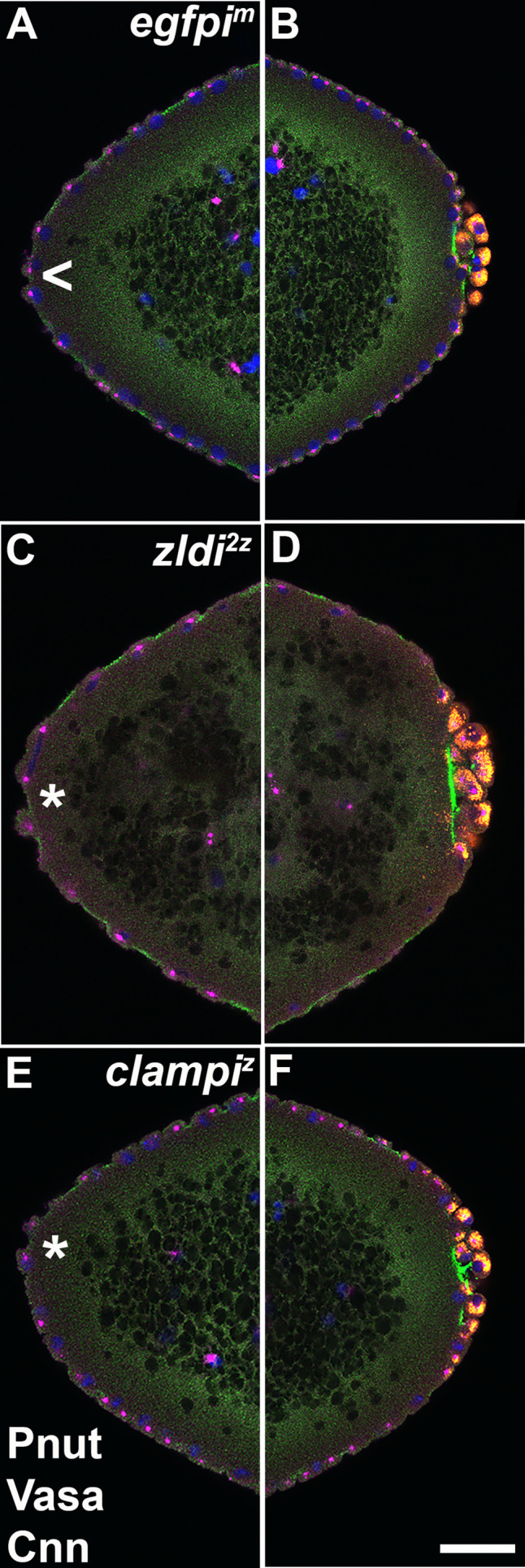
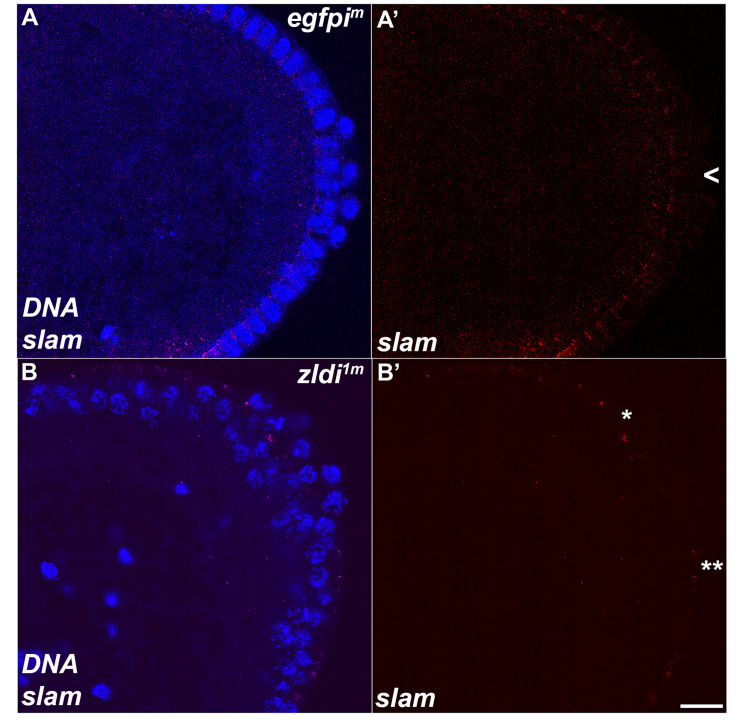
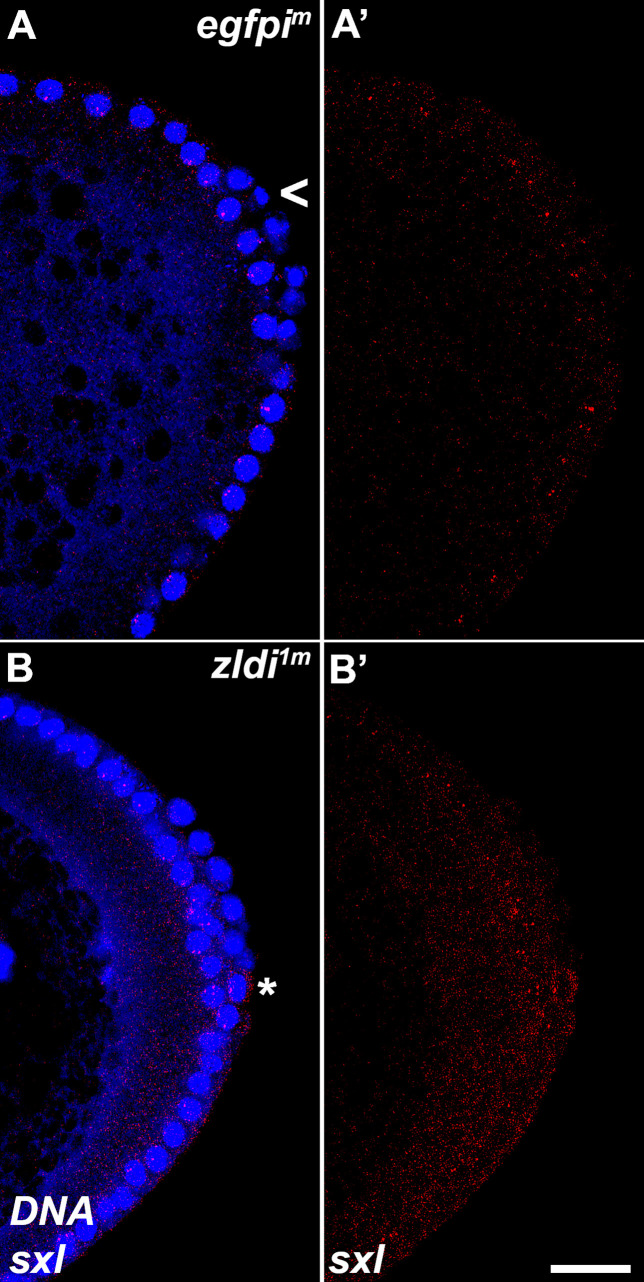
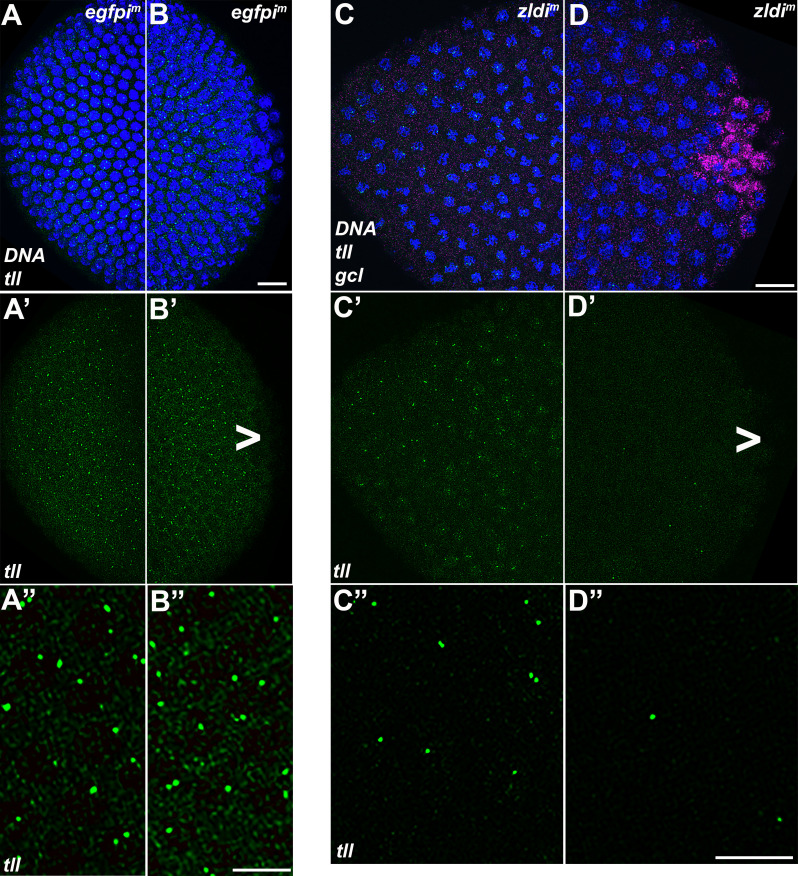
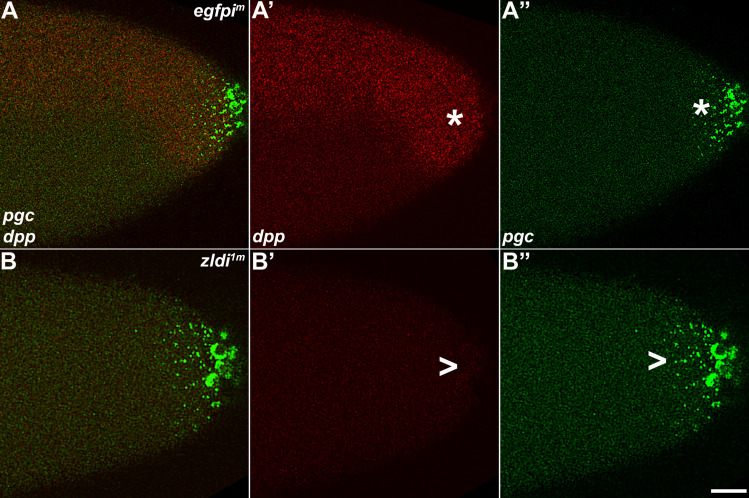
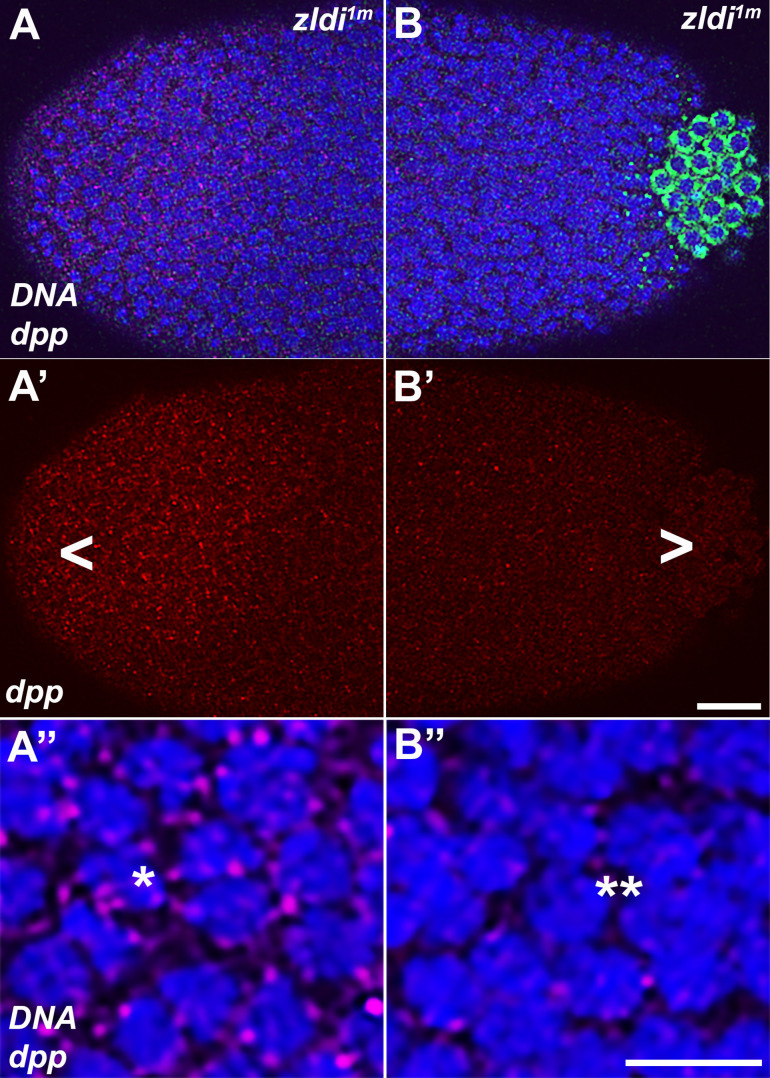
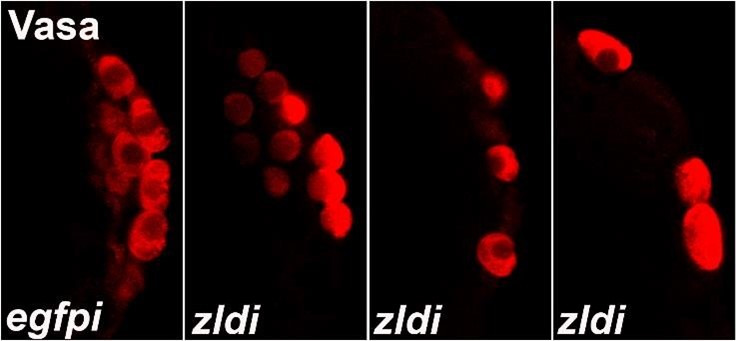
In embryos, somatic versus germline identity is the first cell fate decision. Zygotic genome activation (ZGA) orchestrates regionalized gene expression, imparting specific identity on somatic cells. ZGA begins with a minor wave that commences at nuclear cycle (NC)8 under the guidance of chromatin accessibility factors (Zelda, CLAMP, GAF), followed by the major wave during NC14. By contrast, primordial germ cell (PGC) specification requires maternally deposited and posteriorly anchored germline determinants. This is accomplished by a centrosome coordinated release and sequestration of germ plasm during the precocious cellularization of PGCs in NC10. Here, we report a novel requirement for Zelda and CLAMP during the establishment of the germline/soma distinction. When their activity is compromised, PGC determinants are not properly sequestered, and specification is disrupted. Conversely, the spreading of PGC determinants from the posterior pole adversely influences transcription in the neighboring somatic nuclei. These reciprocal aberrations can be correlated with defects in centrosome duplication/separation that are known to induce inappropriate transmission of the germ plasm. Interestingly, consistent with the ability of bone morphogenetic protein (BMP) signaling to influence specification of embryonic PGCs, reduction in the transcript levels of a BMP family ligand, (), is exacerbated at the posterior pole.
在胚胎中,体细胞核与生殖细胞核的身份是第一个细胞命运决定。合子基因组激活 (ZGA) 协调区域基因表达,赋予体细胞特定的身份。ZGA 始于一个次要波,在染色质可及性因子(Zelda、CLAMP、GAF)的指导下,在核循环 (NC)8 开始,随后在 NC14 期间发生主要波。相比之下,原始生殖细胞 (PGC) 的特化需要母体沉积和后部锚定的生殖系决定因素。这是通过中心体协调释放和隔离生殖质来实现的,在 NC10 中 PGC 的过早细胞化过程中。在这里,我们报告了 Zelda 和 CLAMP 在建立生殖系/体区别中的新要求。当它们的活性受到损害时,PGC 决定因素不能被正确隔离,特化被打乱。相反,PGC 决定因素从后极的扩散会对邻近体细胞核中的转录产生不利影响。这些相互的异常可以与中心体复制/分离的缺陷相关联,已知这些缺陷会导致生殖质的不当传递。有趣的是,与骨形态发生蛋白 (BMP) 信号影响胚胎 PGC 特化的能力一致,BMP 家族配体 ()的转录水平降低在后部极加剧。