Department of Biological Sciences, The University of Texas at El Paso, El Paso, TX, 79968, USA.
Graduate School of Biomedical Sciences, University of Texas Health, San Antonio, TX, 78229, USA.
Sci Rep. 2023 Feb 9;13(1):2309. doi: 10.1038/s41598-023-29357-7.
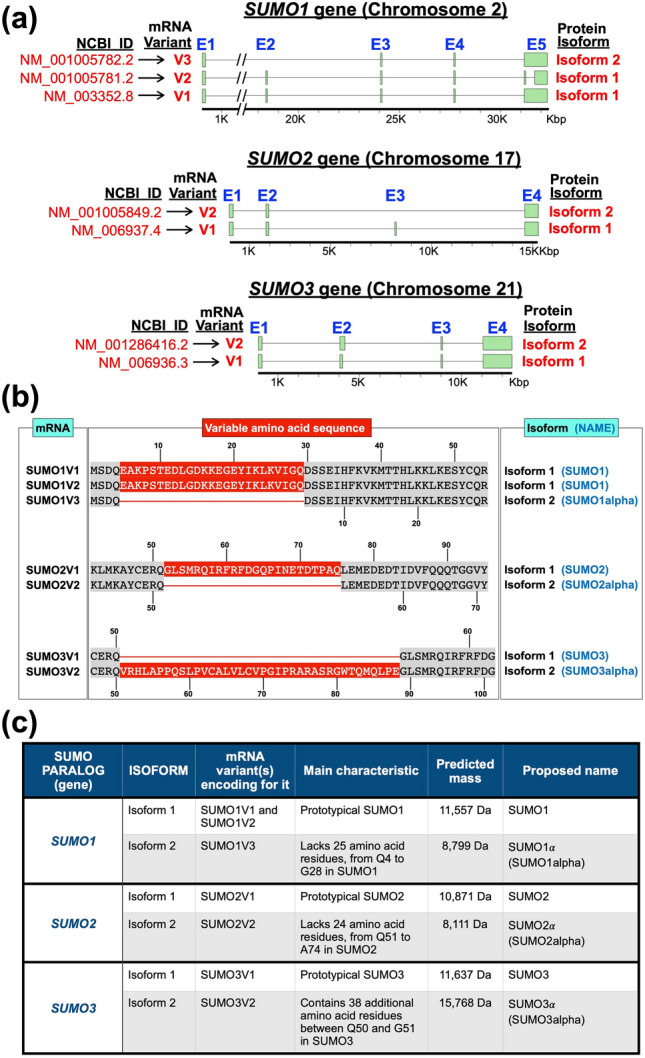
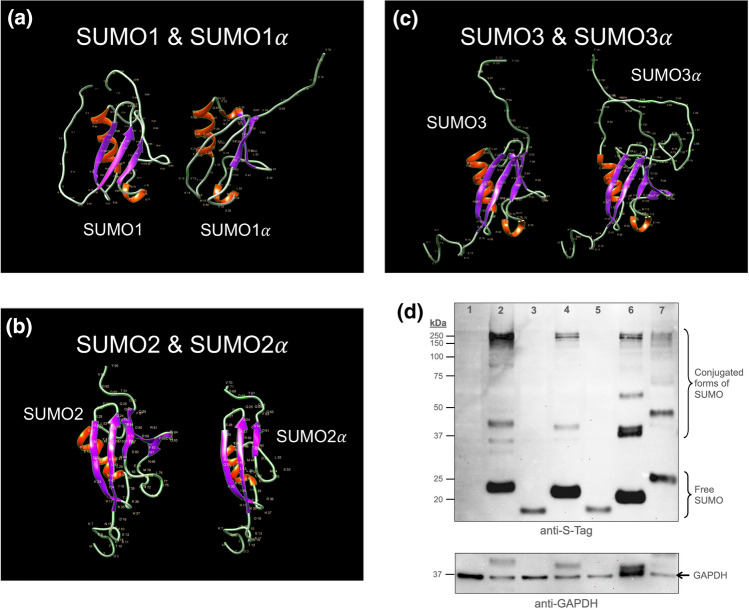
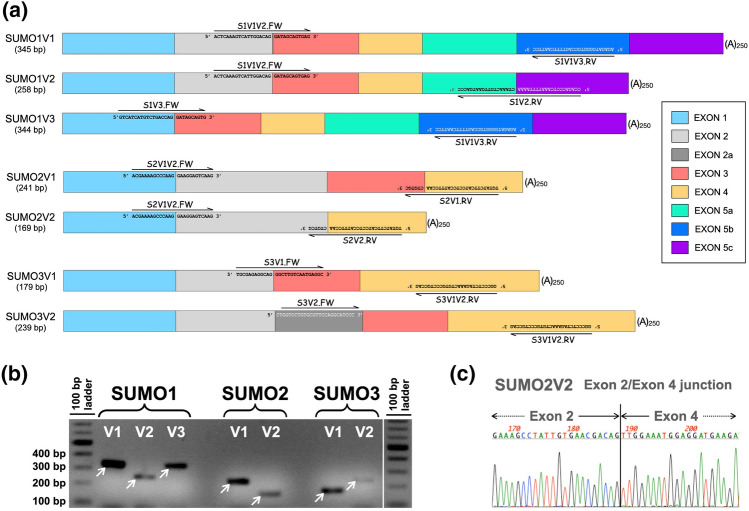
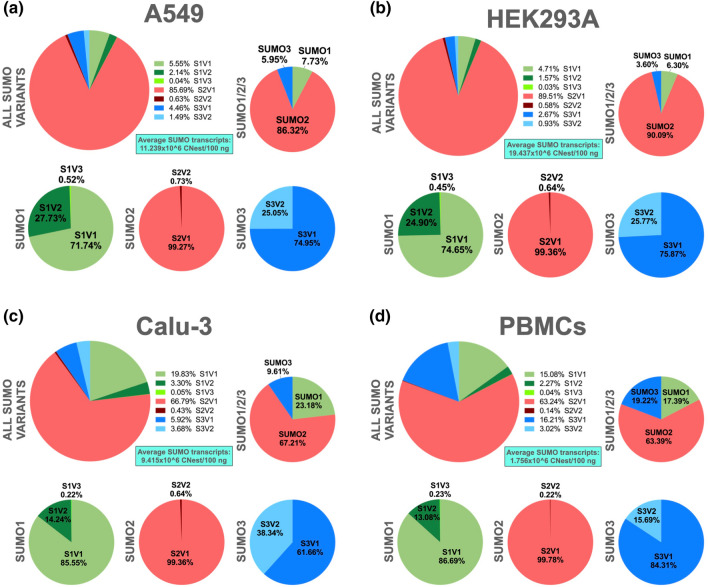
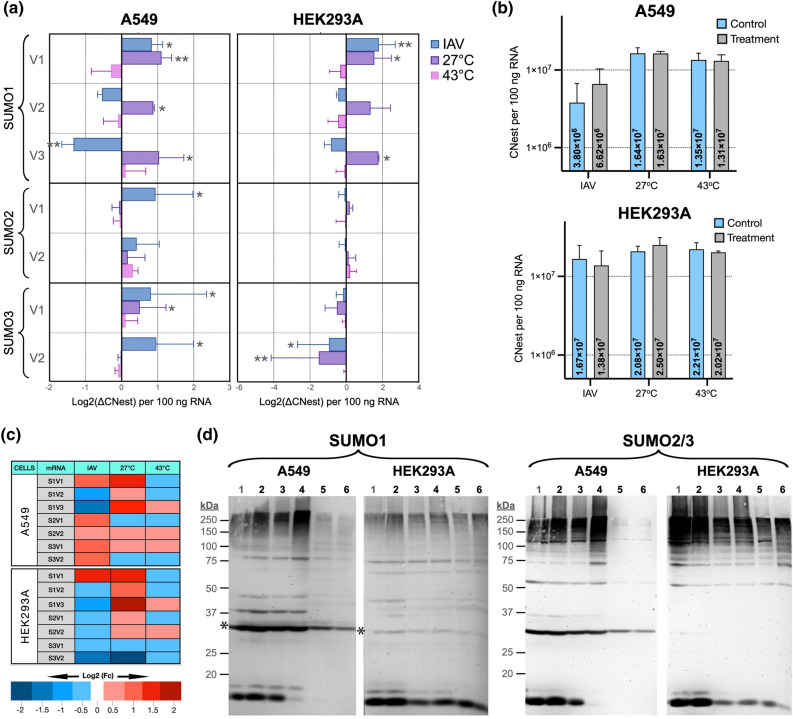
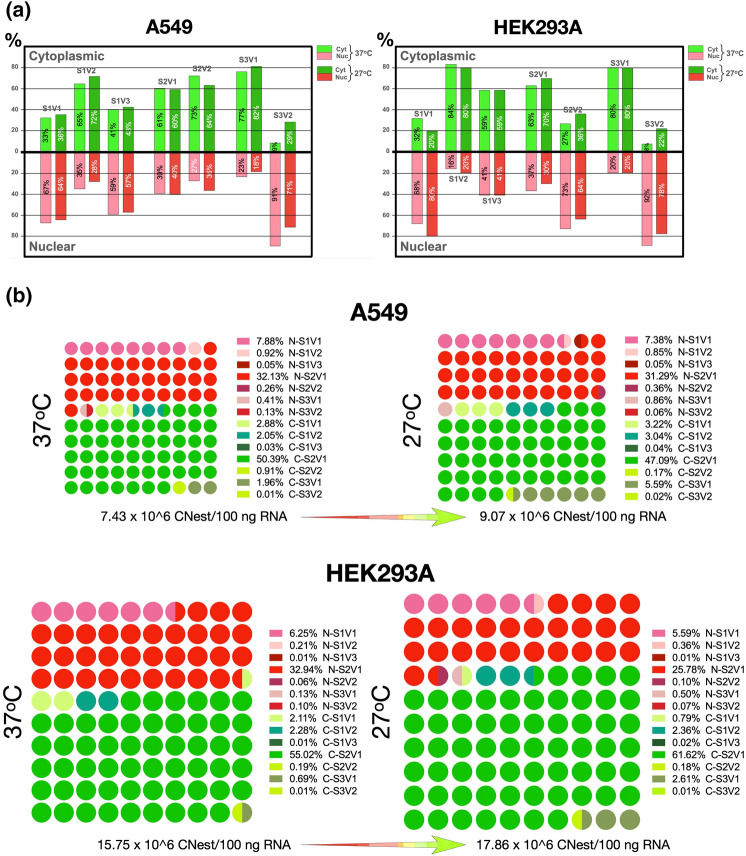
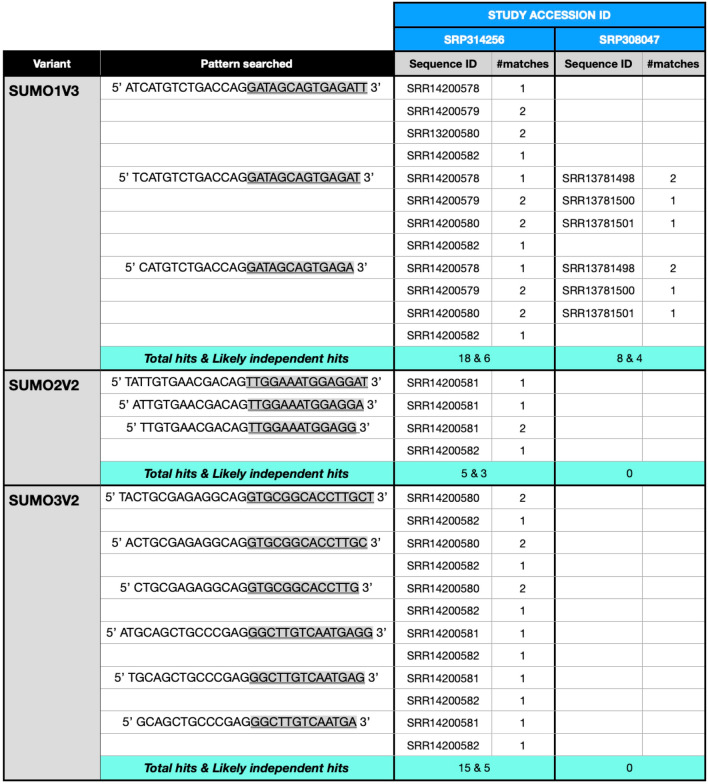
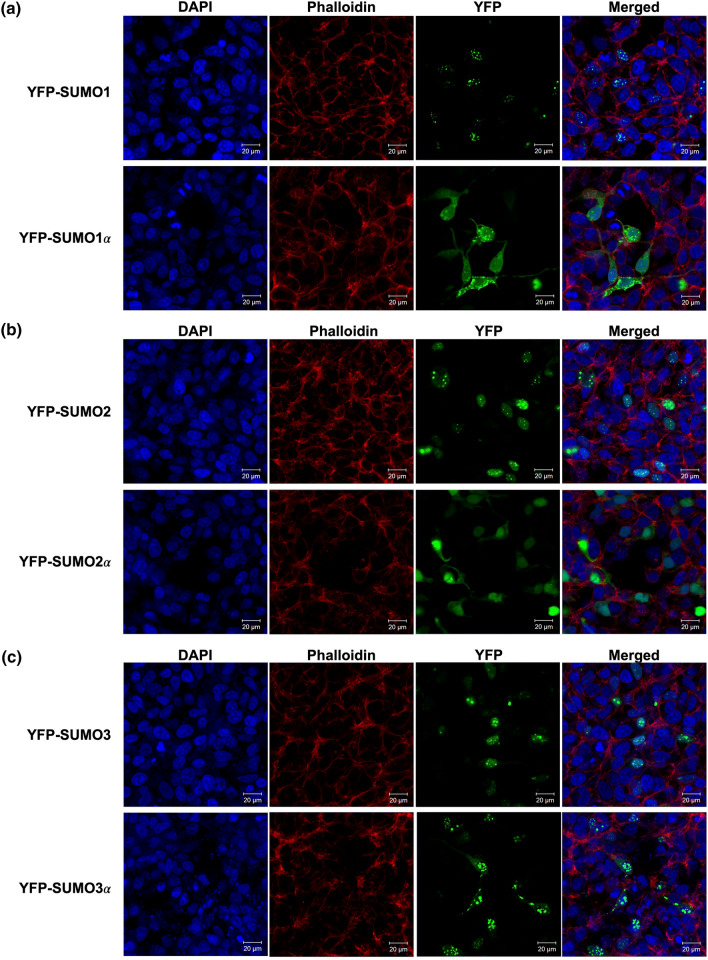
Substantial increases in the conjugation of the main human SUMO paralogs, SUMO1, SUMO2, and SUMO3, are observed upon exposure to different cellular stressors, and such increases are considered important to facilitate cell survival to stress. Despite their critical cellular role, little is known about how the levels of the SUMO modifiers are regulated in the cell, particularly as it relates to the changes observed upon stress. Here we characterize the contribution of alternative splicing towards regulating the expression of the main human SUMO paralogs under normalcy and three different stress conditions, heat-shock, cold-shock, and Influenza A Virus infection. Our data reveal that the normally spliced transcript variants are the predominant mature mRNAs produced from the SUMO genes and that the transcript coding for SUMO2 is by far the most abundant of all. We also provide evidence that alternatively spliced transcripts coding for protein isoforms of the prototypical SUMO proteins, which we refer to as the SUMO alphas, are also produced, and that their abundance and nuclear export are affected by stress in a stress- and cell-specific manner. Additionally, we provide evidence that the SUMO alphas are actively synthesized in the cell as their coding mRNAs are found associated with translating ribosomes. Finally, we provide evidence that the SUMO alphas are functionally different from their prototypical counterparts, with SUMO1α and SUMO2α being non-conjugatable to protein targets, SUMO3α being conjugatable but targeting a seemingly different subset of protein from those targeted by SUMO3, and all three SUMO alphas displaying different cellular distributions from those of the prototypical SUMOs. Thus, alternative splicing appears to be an important contributor to the regulation of the expression of the SUMO proteins and the cellular functions of the SUMOylation system.
在暴露于不同的细胞应激源时,主要的人类 SUMO 同工型 SUMO1、SUMO2 和 SUMO3 的缀合显著增加,并且这种增加被认为对于促进细胞对应激的存活很重要。尽管它们在细胞中具有关键作用,但对于 SUMO 修饰物水平如何在细胞中进行调节,特别是与应激时观察到的变化相关的调节,知之甚少。在这里,我们描述了在正常情况下和三种不同应激条件下,热休克、冷休克和流感 A 病毒感染,替代剪接对主要人类 SUMO 同工型表达的调控作用。我们的数据表明,正常剪接的转录变体是从 SUMO 基因产生的主要成熟 mRNA,并且编码 SUMO2 的转录本是所有转录本中最丰富的。我们还提供了证据表明,编码典型 SUMO 蛋白的蛋白同工型的替代剪接转录本也被产生,并且它们的丰度和核输出受到应激的影响,具有应激和细胞特异性。此外,我们提供了证据表明 SUMO 阿尔法是在细胞中主动合成的,因为它们的编码 mRNA 与翻译核糖体相关联。最后,我们提供了证据表明 SUMO 阿尔法与它们的原型对应物在功能上不同,SUMO1α 和 SUMO2α 不能与蛋白靶标缀合,SUMO3α 可以缀合但靶向的蛋白似乎与 SUMO3 靶向的蛋白不同,并且所有三个 SUMO 阿尔法都表现出与原型 SUMO 不同的细胞分布。因此,替代剪接似乎是调节 SUMO 蛋白表达和 SUMO 化系统细胞功能的重要因素。