Institute of Cardiovascular Organogenesis and Regeneration, Faculty of Medicine, WWU Münster, Münster, Germany.
Münster Imaging Network, Cells in Motion Interfaculty Centre, Faculty of Biology, WWU Münster, Münster, Germany.
Elife. 2023 Apr 25;12:e82969. doi: 10.7554/eLife.82969.
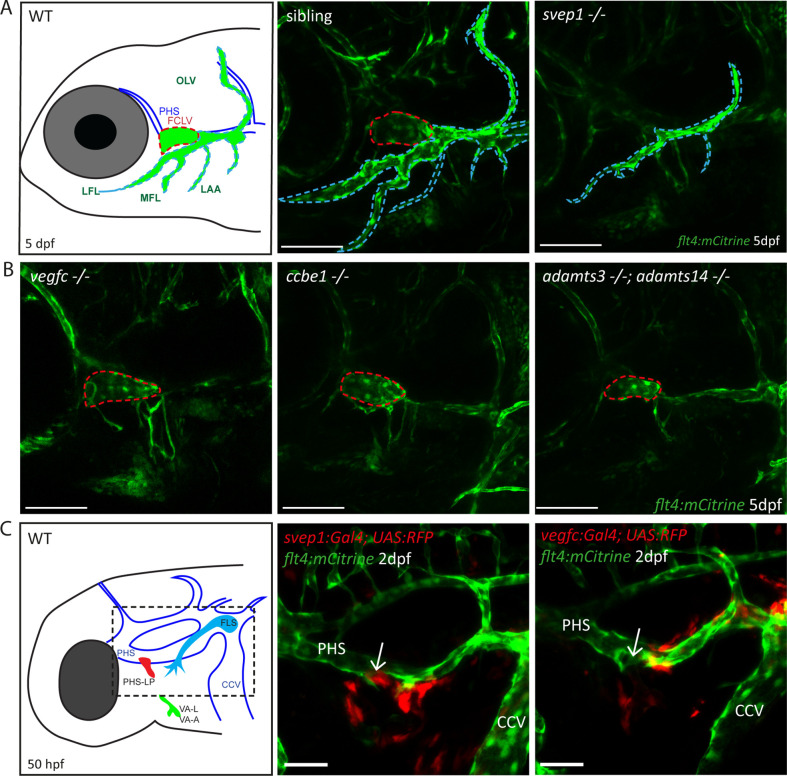
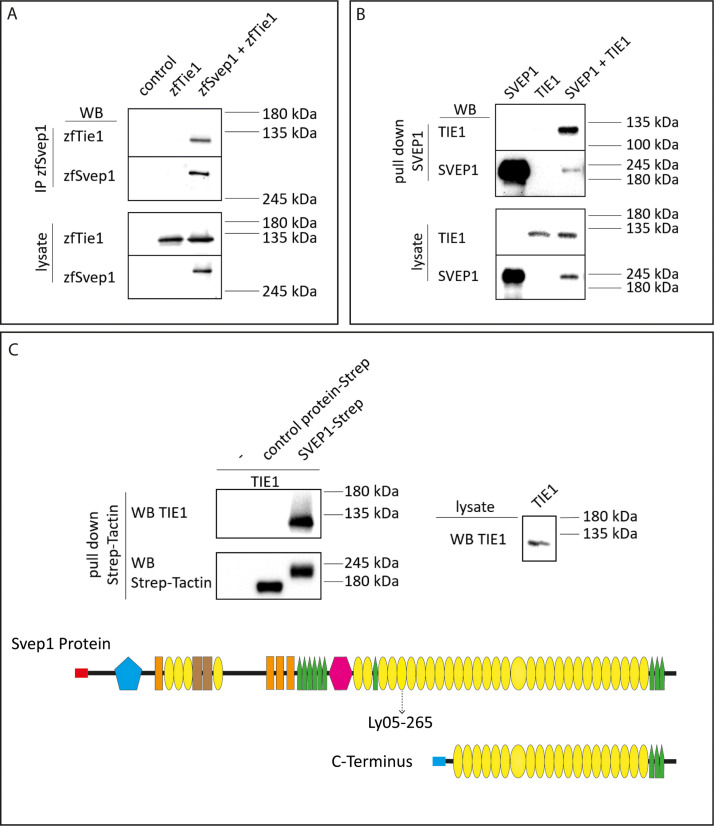
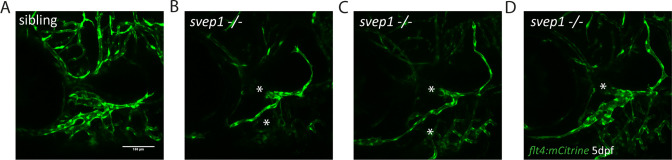
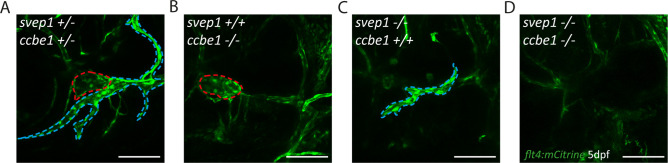
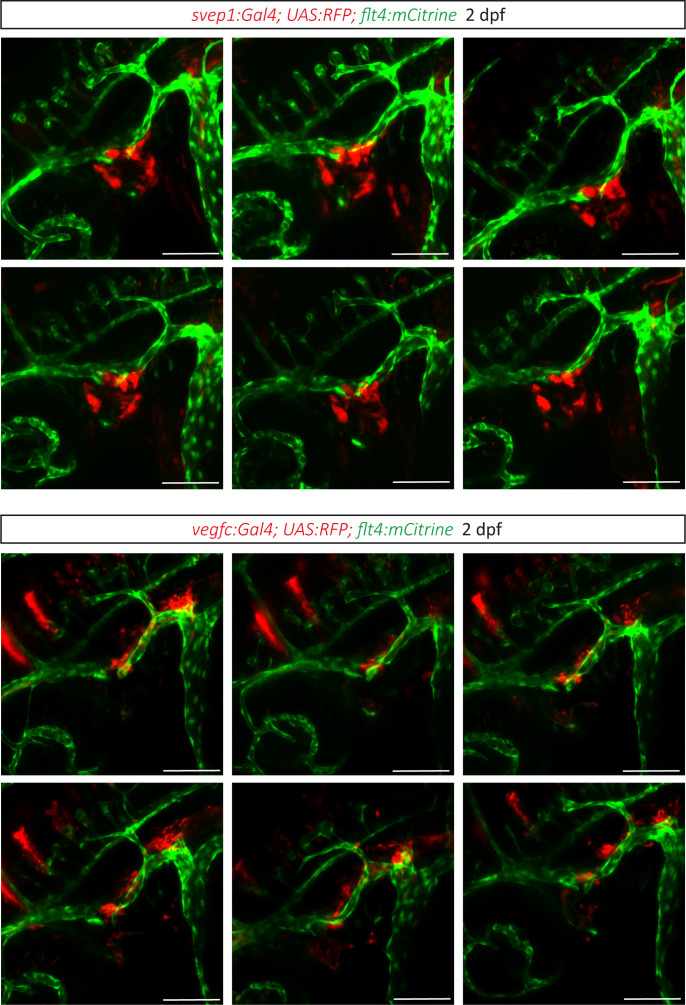
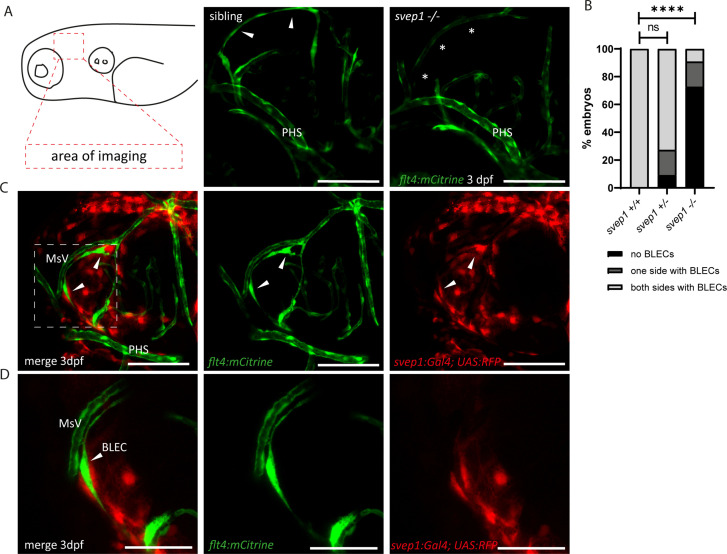
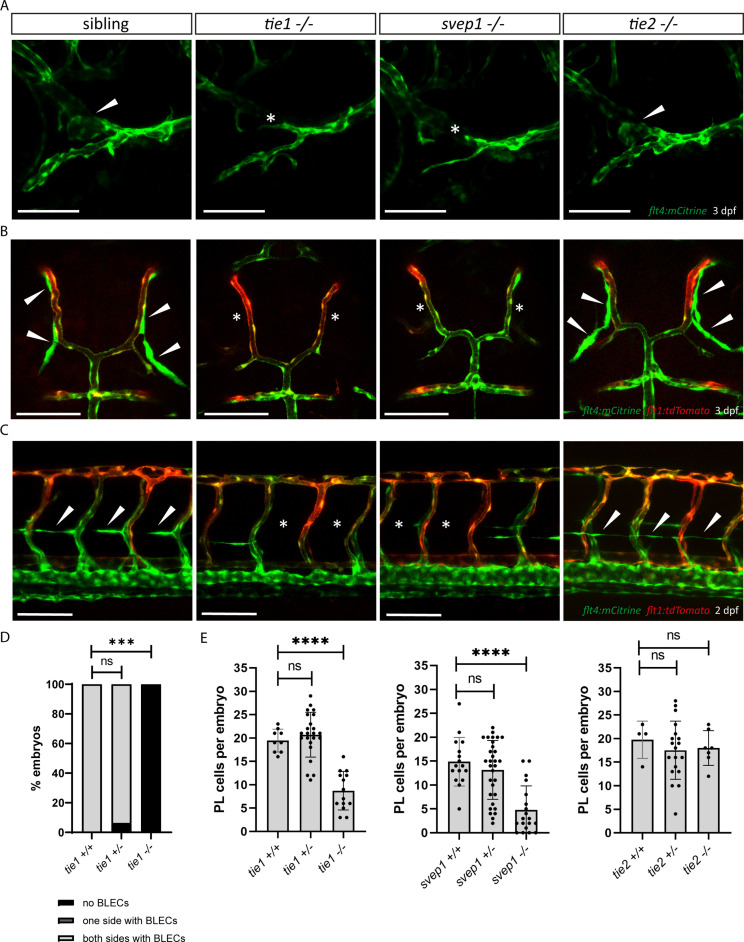
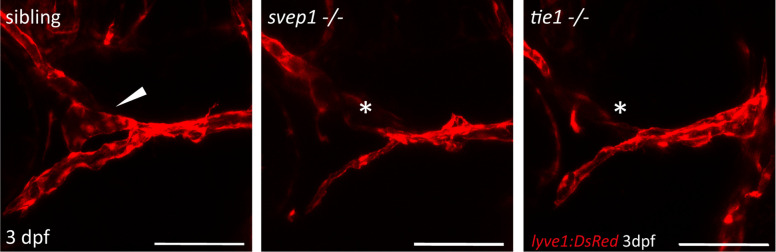
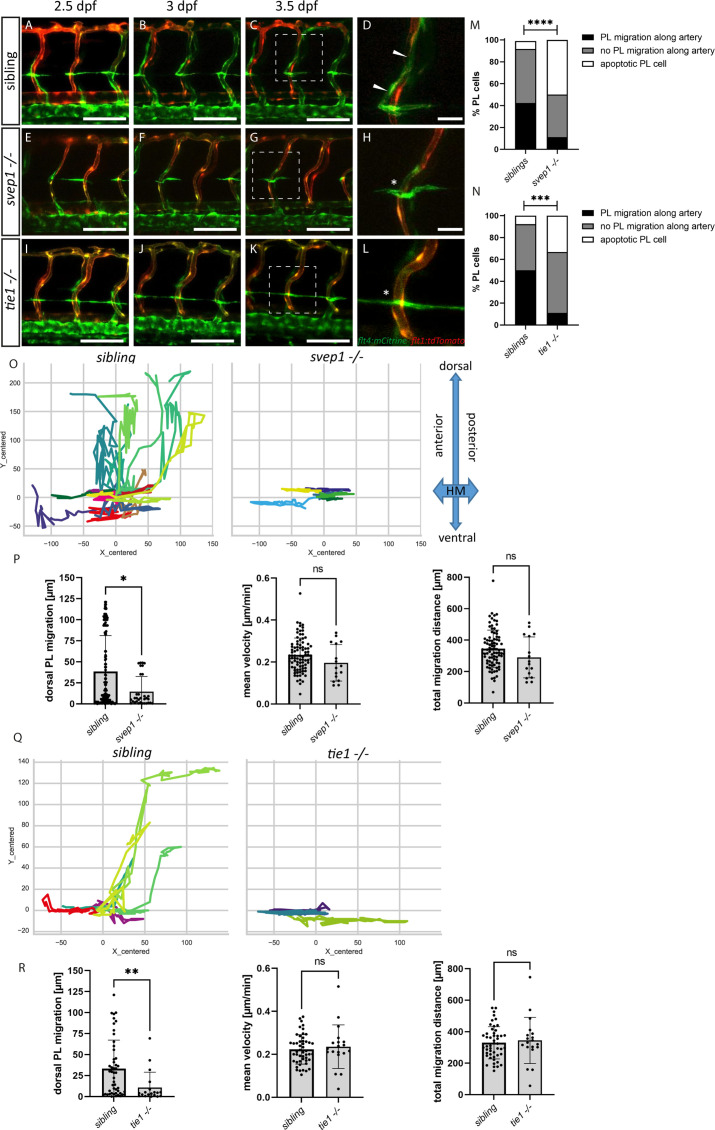
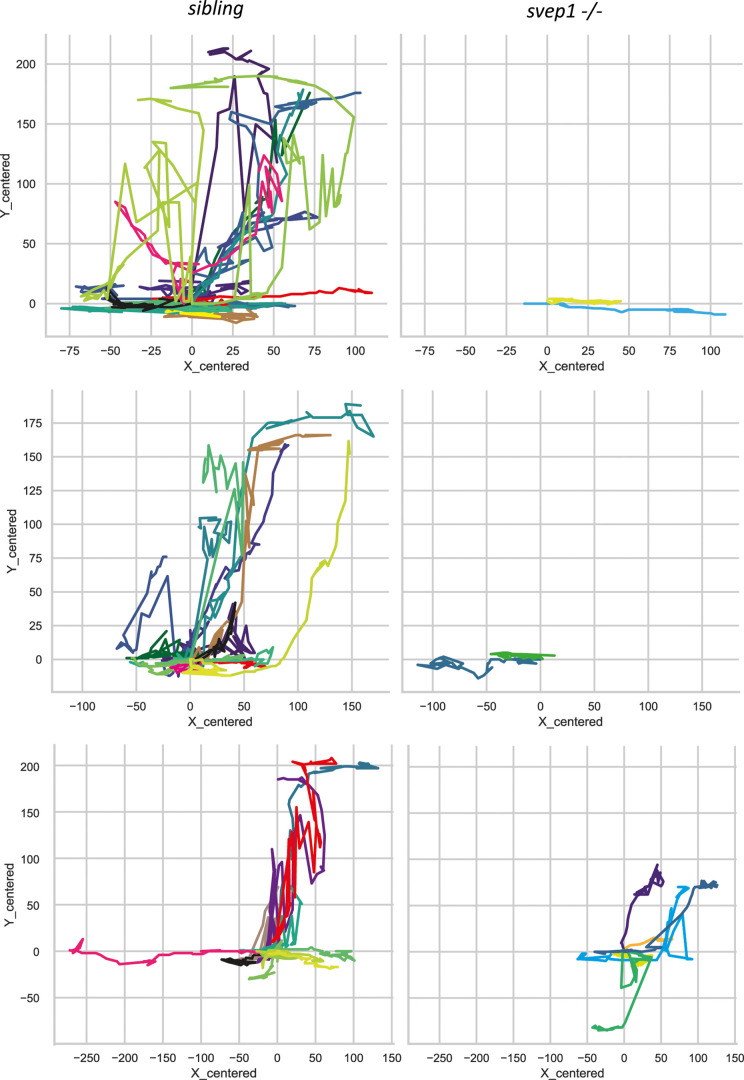
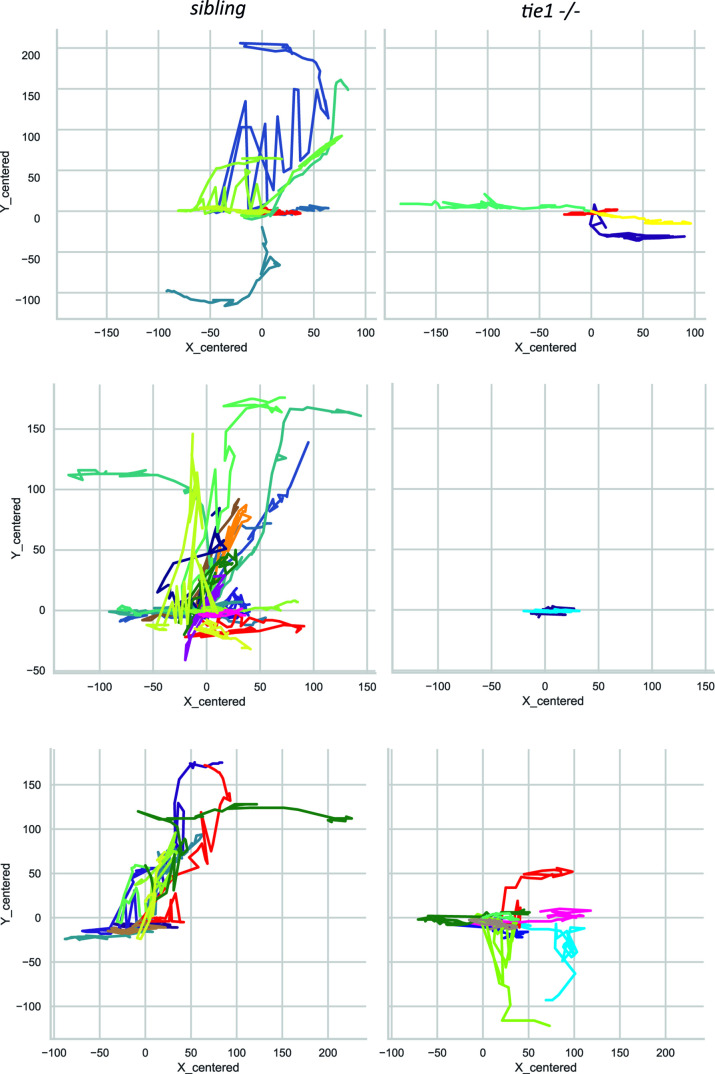
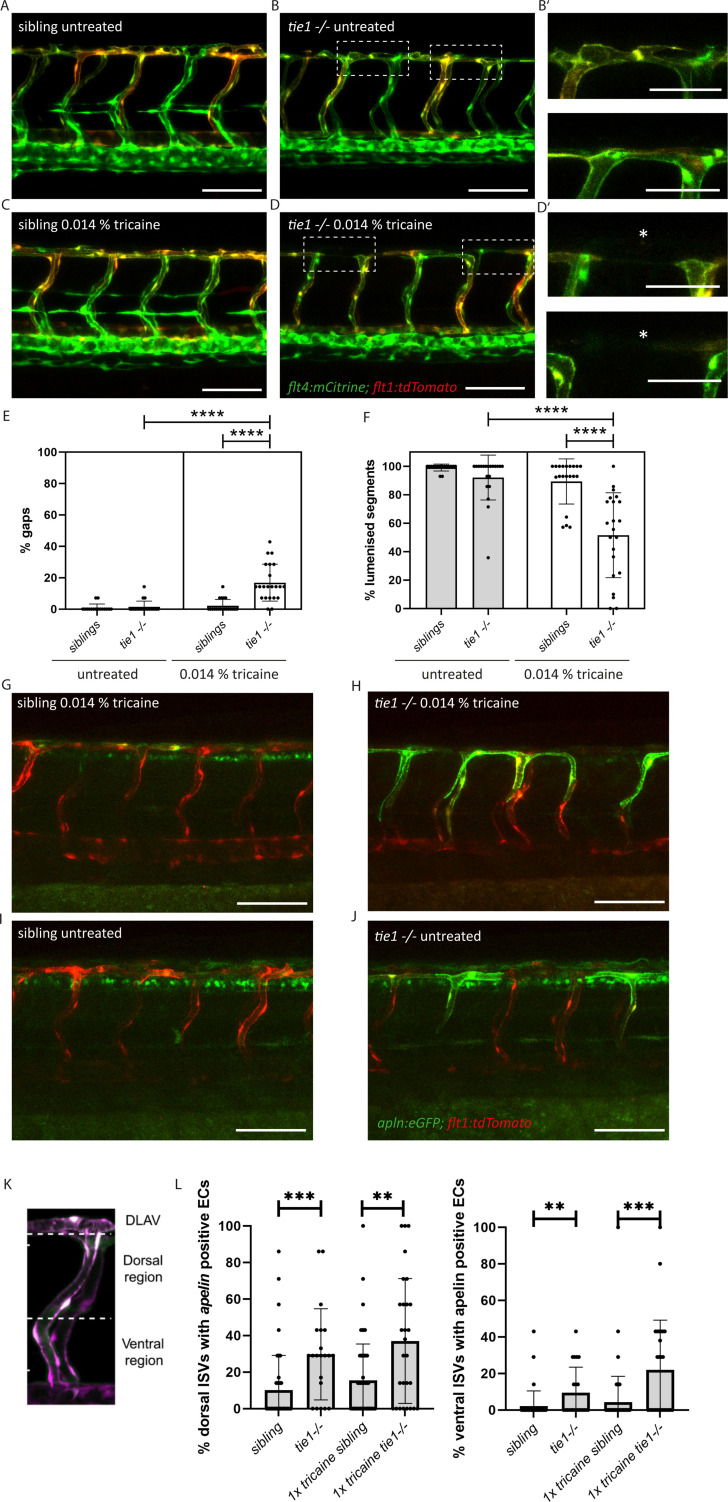
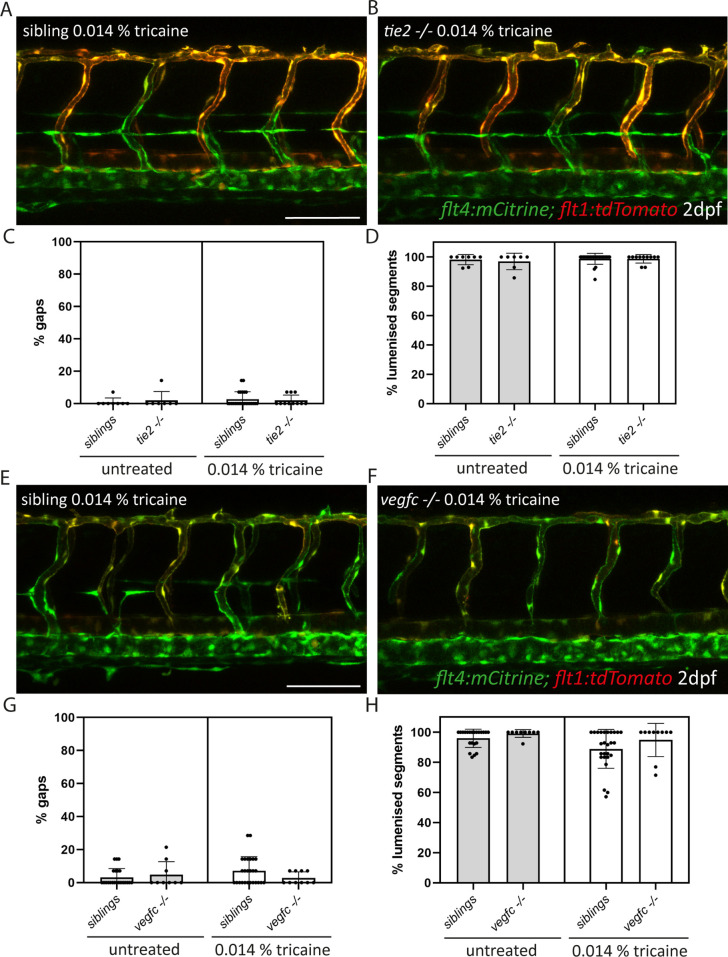
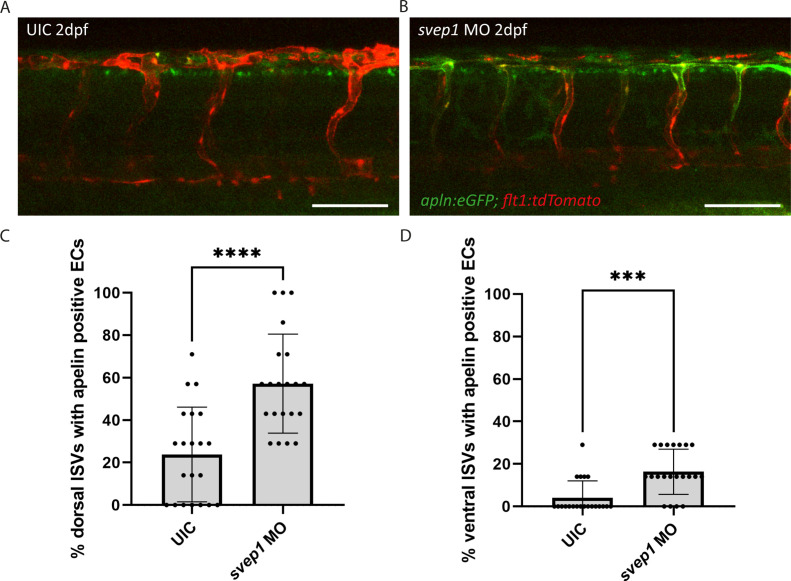
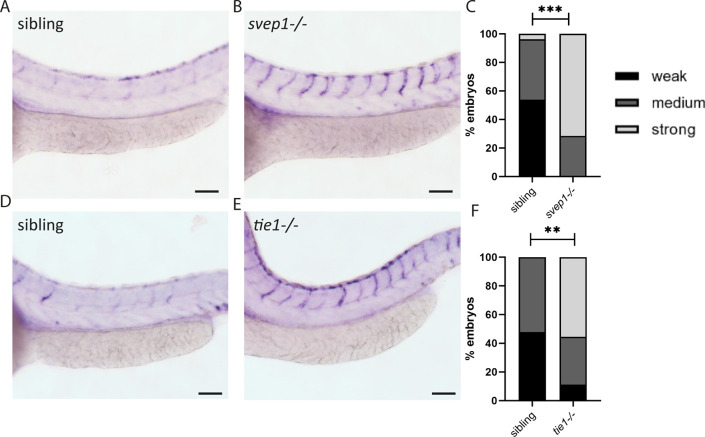
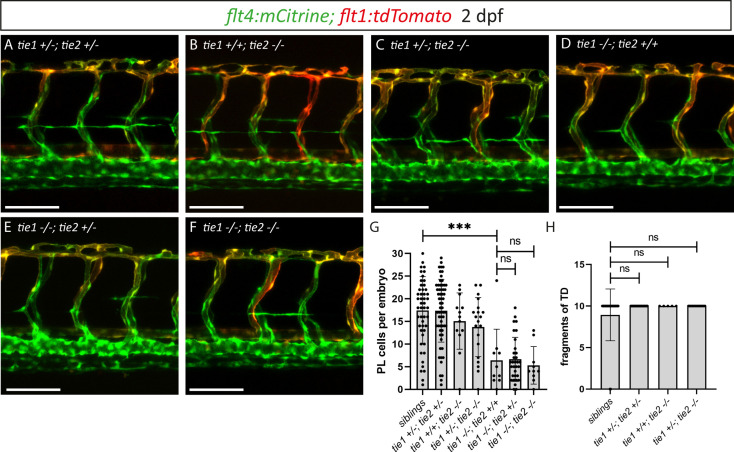
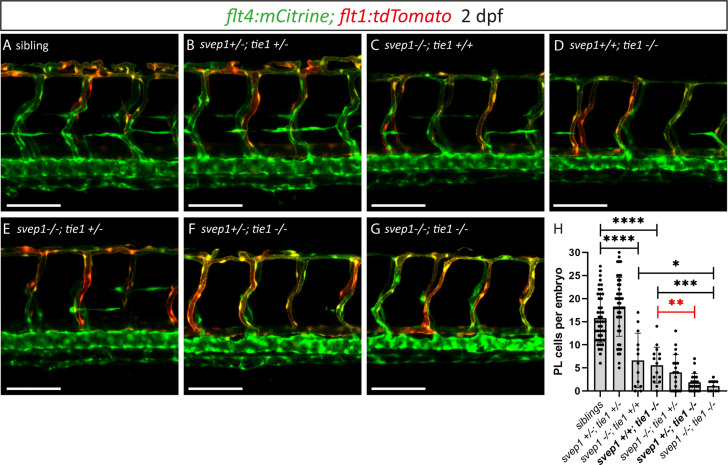
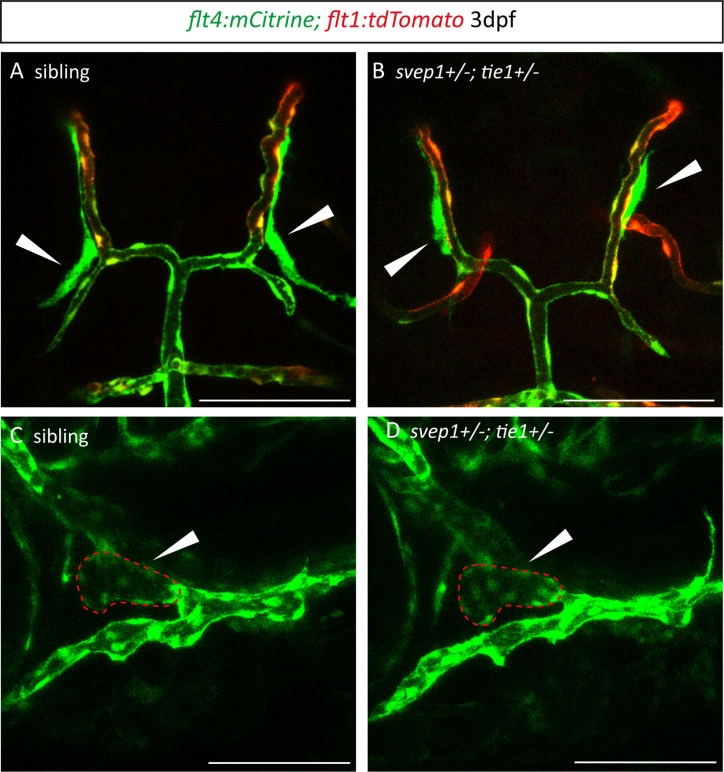
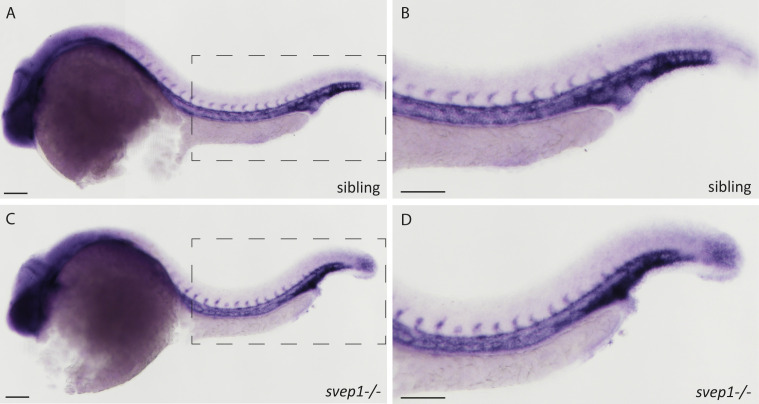
Multiple factors are required to form functional lymphatic vessels. Here, we uncover an essential role for the secreted protein Svep1 and the transmembrane receptor Tie1 during the development of subpopulations of the zebrafish facial lymphatic network. This specific aspect of the facial network forms independently of Vascular endothelial growth factor C (Vegfc) signalling, which otherwise is the most prominent signalling axis in all other lymphatic beds. Additionally, we find that multiple specific and newly uncovered phenotypic hallmarks of mutants are also present in , but not in or mutants. These phenotypes are observed in the lymphatic vasculature of both head and trunk, as well as in the development of the dorsal longitudinal anastomotic vessel under reduced flow conditions. Therefore, our study demonstrates an important function for Tie1 signalling during lymphangiogenesis as well as blood vessel development in zebrafish. Furthermore, we show genetic interaction between and in vivo, during early steps of lymphangiogenesis, and demonstrate that zebrafish as well as human Svep1/SVEP1 protein bind to the respective Tie1/TIE1 receptors in vitro. Since compound heterozygous mutations for and have recently been reported in human glaucoma patients, our data have clinical relevance in demonstrating a role for SVEP1 in TIE signalling in an in vivo setting.
多种因素是形成功能性淋巴管所必需的。在这里,我们揭示了分泌蛋白 Svep1 和跨膜受体 Tie1 在斑马鱼面部淋巴管网络亚群发育过程中的重要作用。面部网络的这一特定方面的形成独立于血管内皮生长因子 C(Vegfc)信号,而 Vegfc 信号是所有其他淋巴管床中最突出的信号轴。此外,我们发现, 突变体的多个特定和新发现的表型特征也存在于 突变体中,但不存在于 或 突变体中。这些表型在头部和躯干的淋巴管血管以及在减少血流条件下背侧纵向吻合血管的发育中都可以观察到。因此,我们的研究表明 Tie1 信号在斑马鱼淋巴管生成以及血管发育过程中具有重要功能。此外,我们还在体内早期的淋巴管生成过程中显示了 和 之间的遗传相互作用,并证明了斑马鱼和人类 Svep1/SVEP1 蛋白在体外与相应的 Tie1/TIE1 受体结合。由于 和 的复合杂合突变最近在人类青光眼患者中被报道,我们的数据在证明 SVEP1 在体内 TIE 信号中的作用方面具有临床相关性。