Centre for Research in Agricultural Genomics, CRAG (CSIC-IRTA-UAB-UB), Campus UAB, Cerdanyola del Vallès, Barcelona, Spain.
Center for Genomics and Systems Biology, New York University, New York, United States.
Elife. 2023 Jul 19;12:RP86324. doi: 10.7554/eLife.86324.
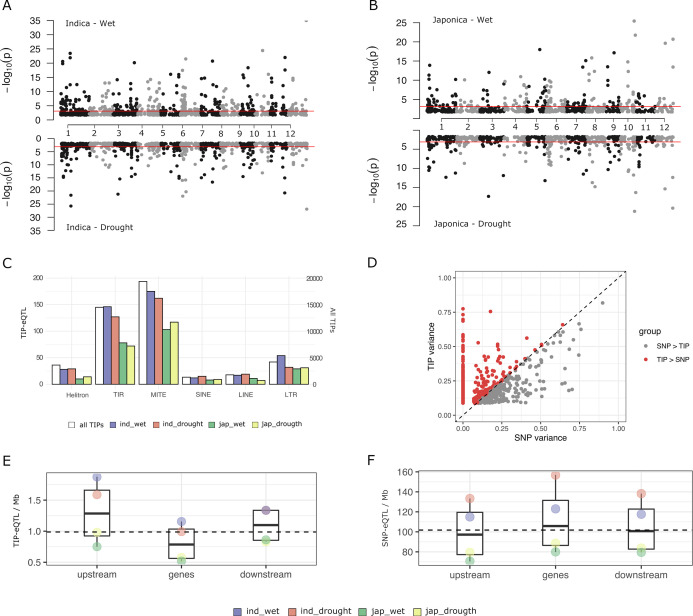
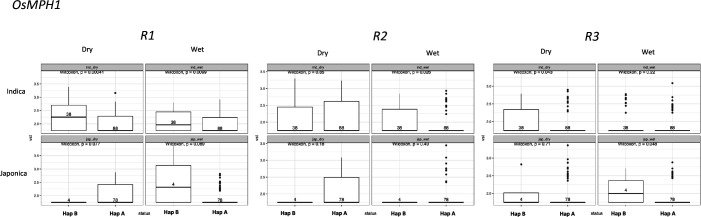
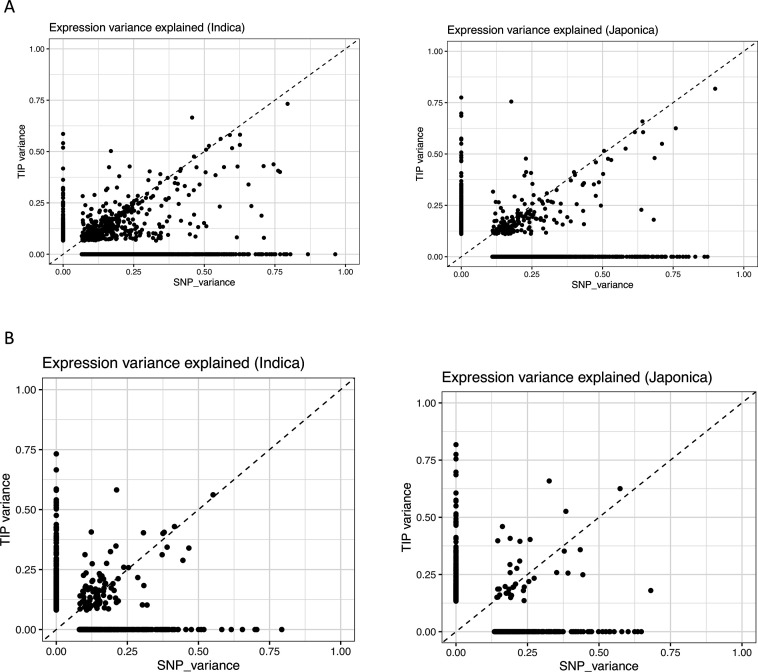
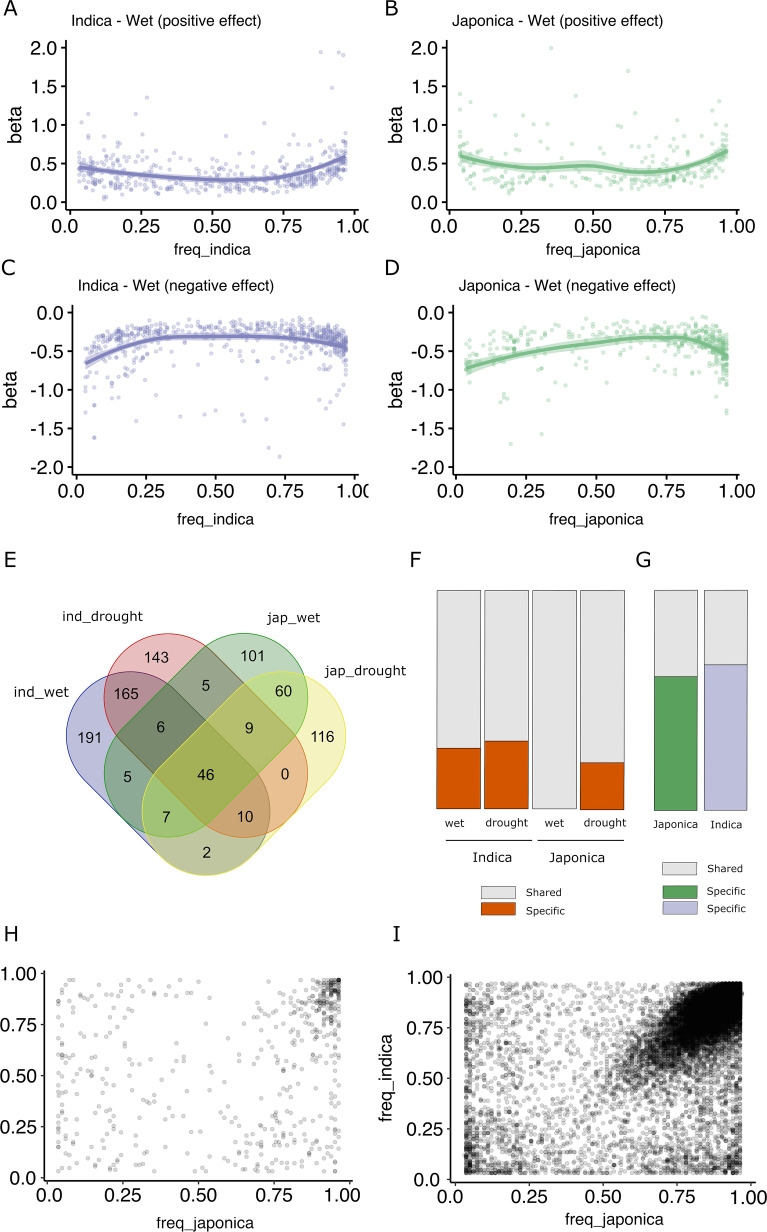
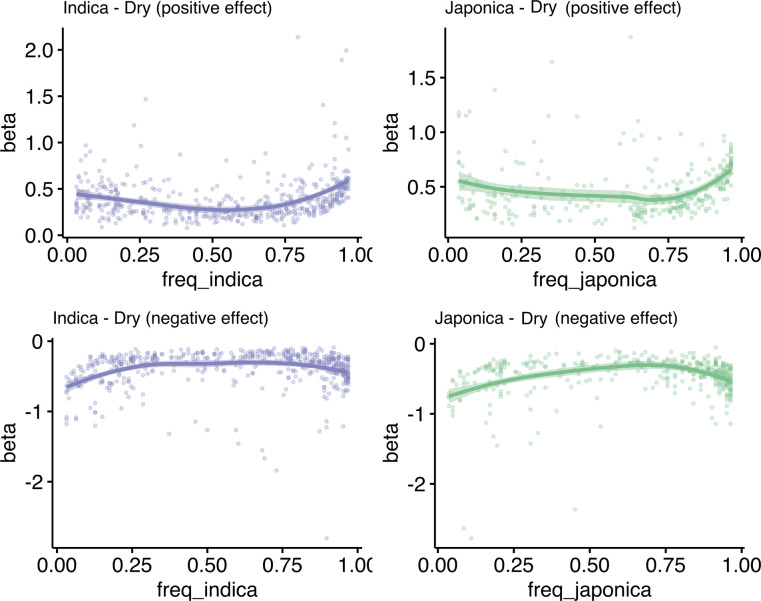
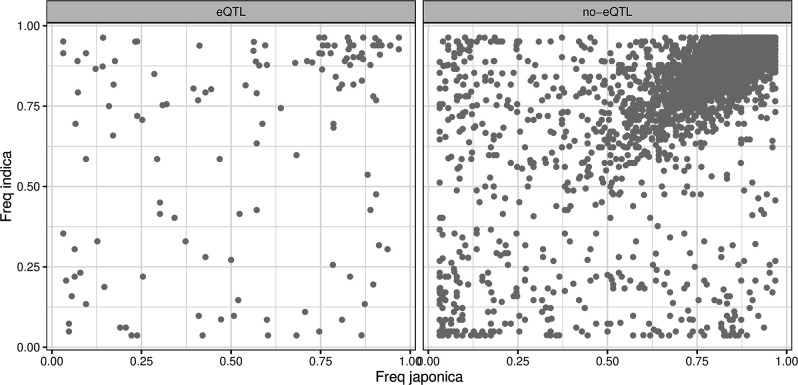
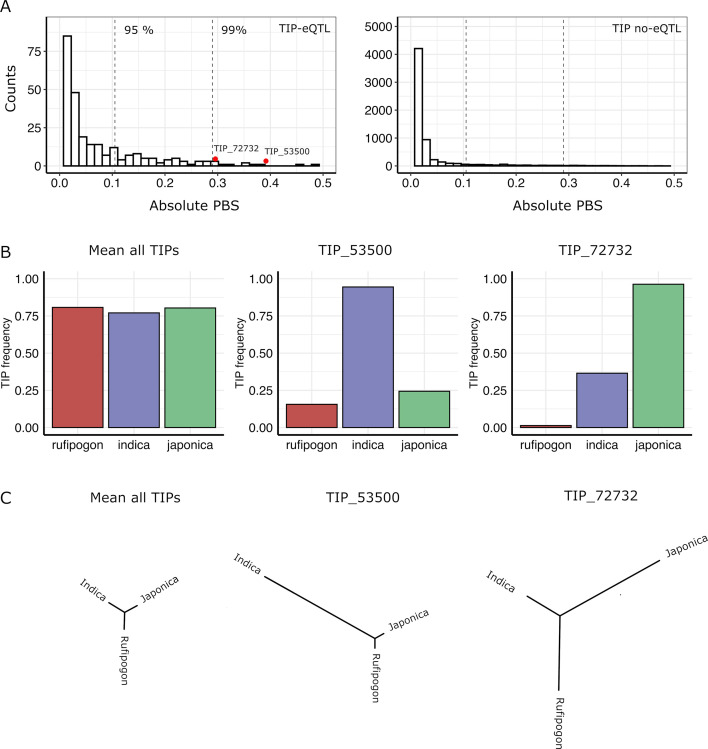
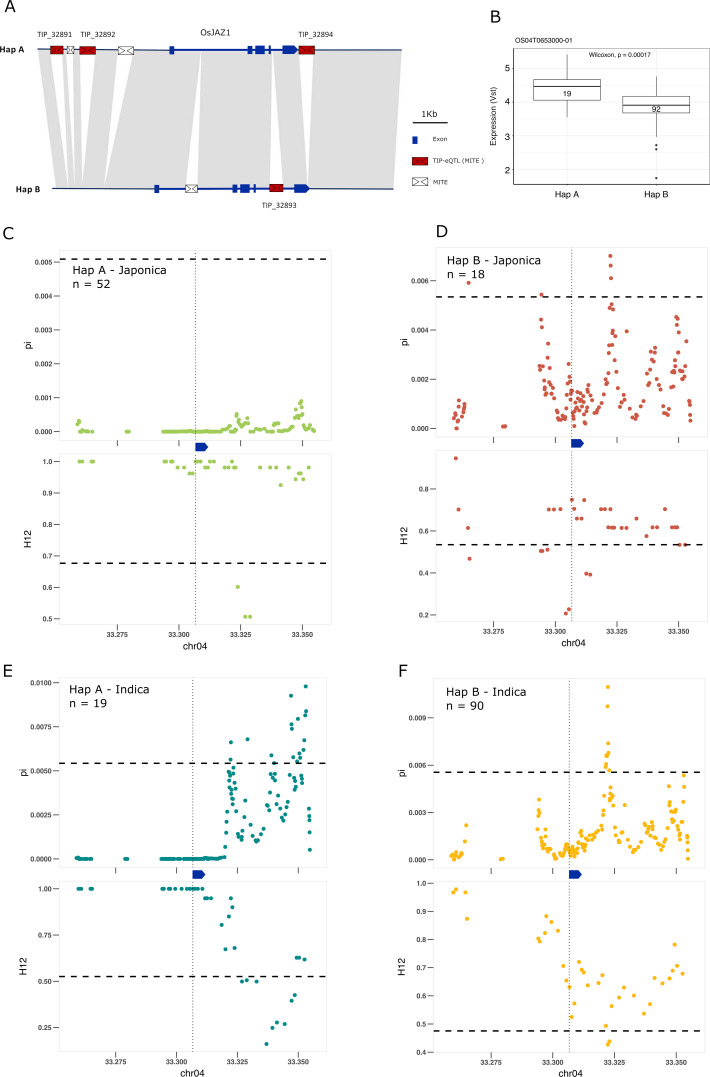
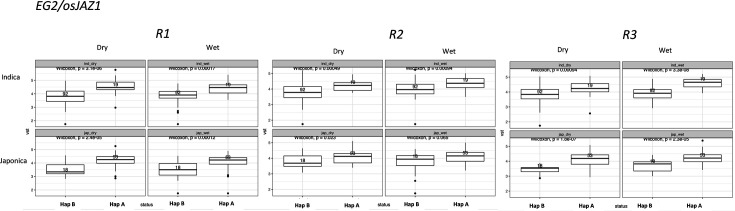
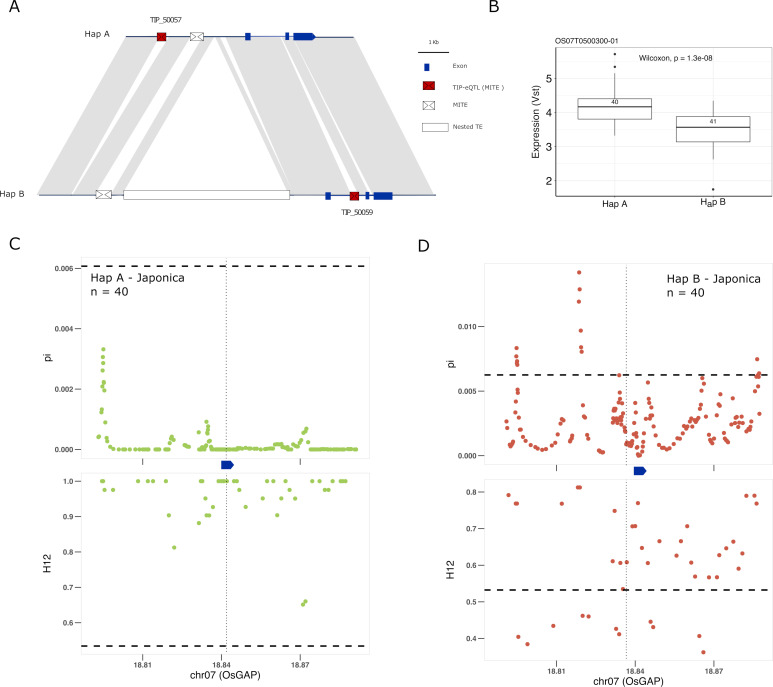
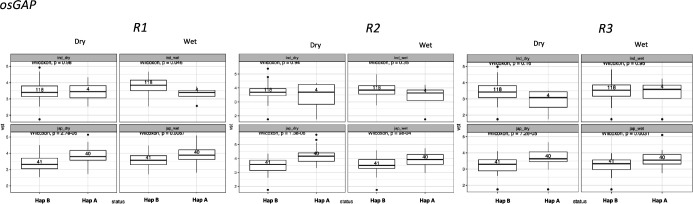
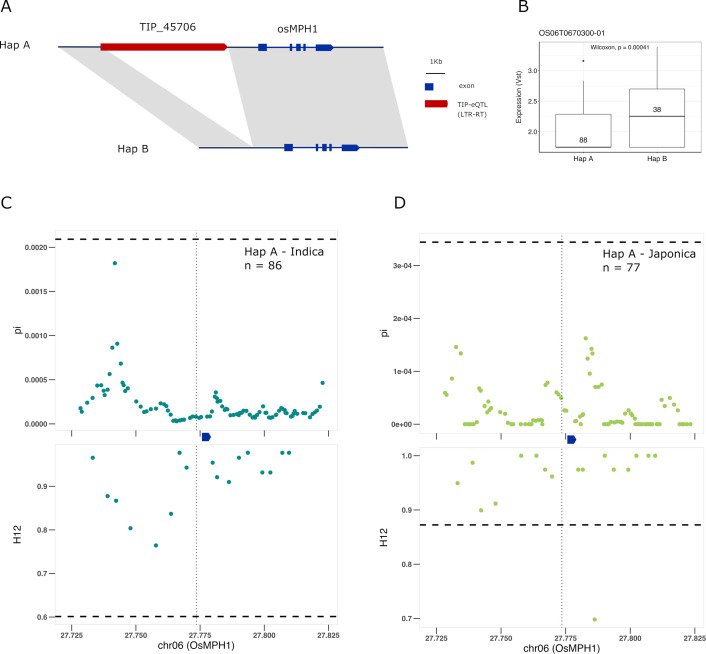
Transposable elements (TEs) are an important source of genome variability. Here, we analyze their contribution to gene expression variability in rice by performing a TE insertion polymorphism expression quantitative trait locus mapping using expression data from 208 varieties from the ssp. and ssp. subspecies. Our data show that TE insertions are associated with changes of expression of many genes known to be targets of rice domestication and breeding. An important fraction of these insertions were already present in the rice wild ancestors, and have been differentially selected in indica and japonica rice populations. Taken together, our results show that small changes of expression in signal transduction genes induced by TE insertions accompany the domestication and adaptation of rice populations.
转座元件(TEs)是基因组变异性的重要来源。在这里,我们通过对来自 亚种和 亚种的 208 个品种的表达数据进行 TE 插入多态性表达数量性状位点作图,分析了它们对水稻基因表达变异性的贡献。我们的数据表明,TE 插入与许多已知是水稻驯化和选育目标的基因表达变化有关。这些插入中有很大一部分已经存在于水稻野生祖先中,并在籼稻和粳稻群体中被差异选择。总的来说,我们的结果表明,TE 插入诱导的信号转导基因表达的微小变化伴随着水稻种群的驯化和适应。