Oxford Protein Informatics Group, Department of Statistics, University of Oxford, Oxford, United Kingdom.
Twist Bioscience, South San Francisco, CA, United States.
Front Immunol. 2023 Jul 18;14:1231623. doi: 10.3389/fimmu.2023.1231623. eCollection 2023.
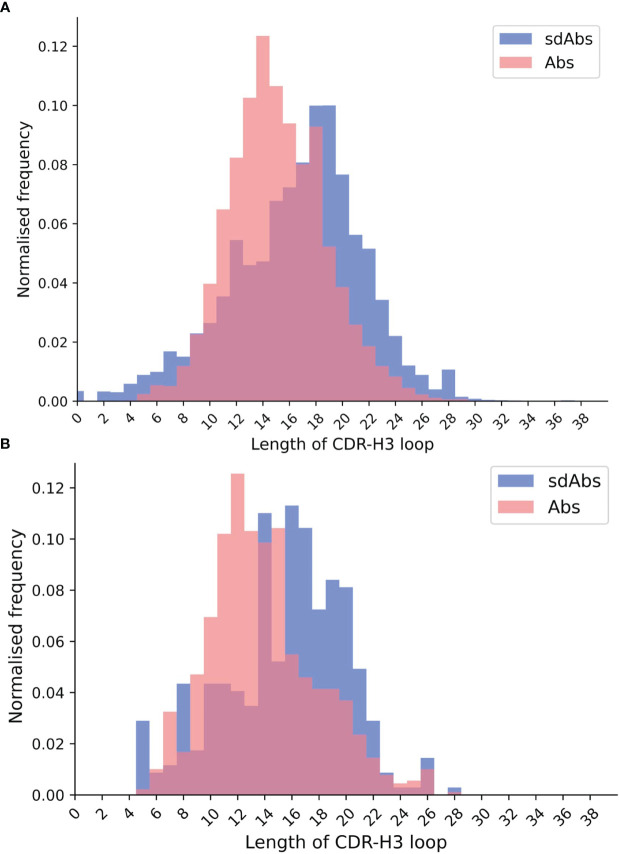
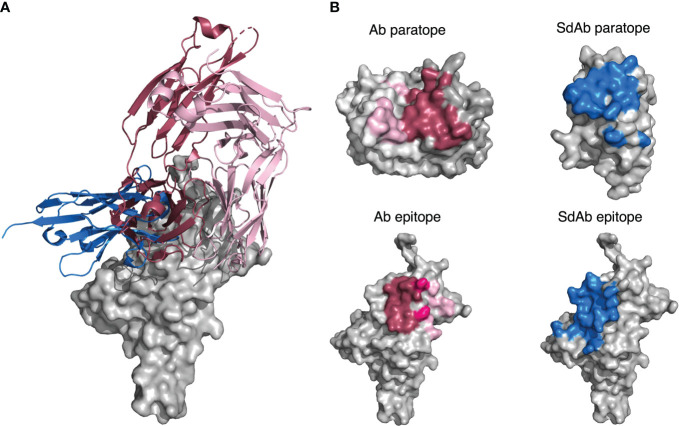
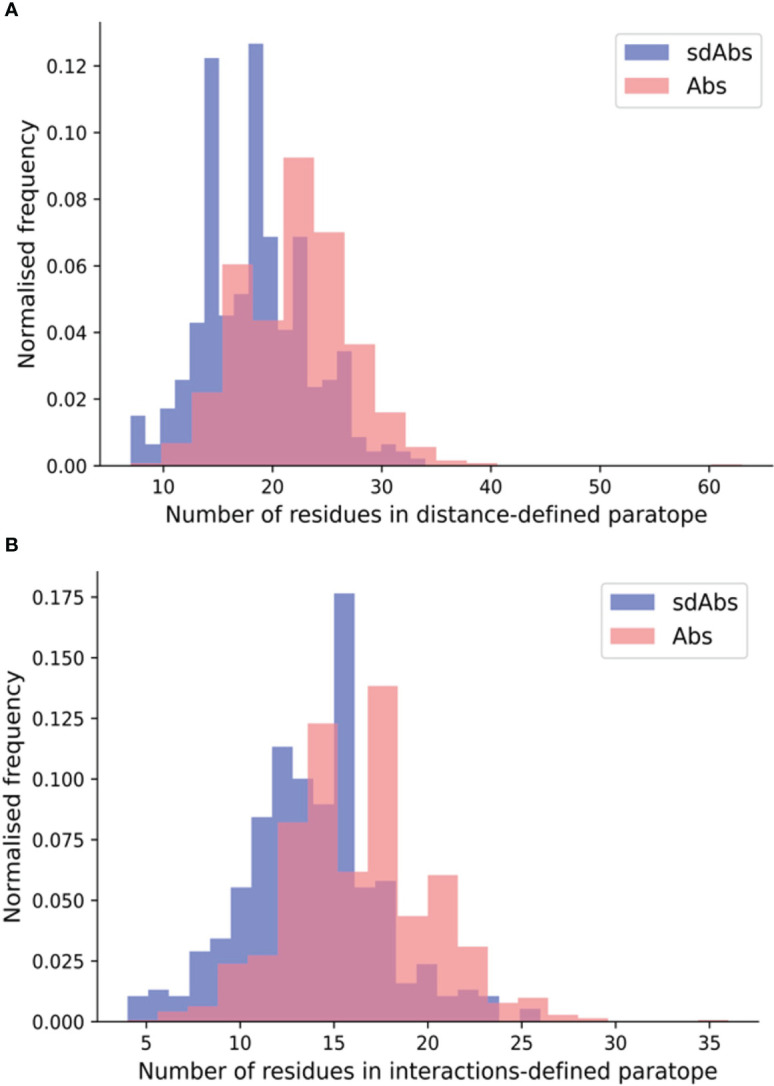
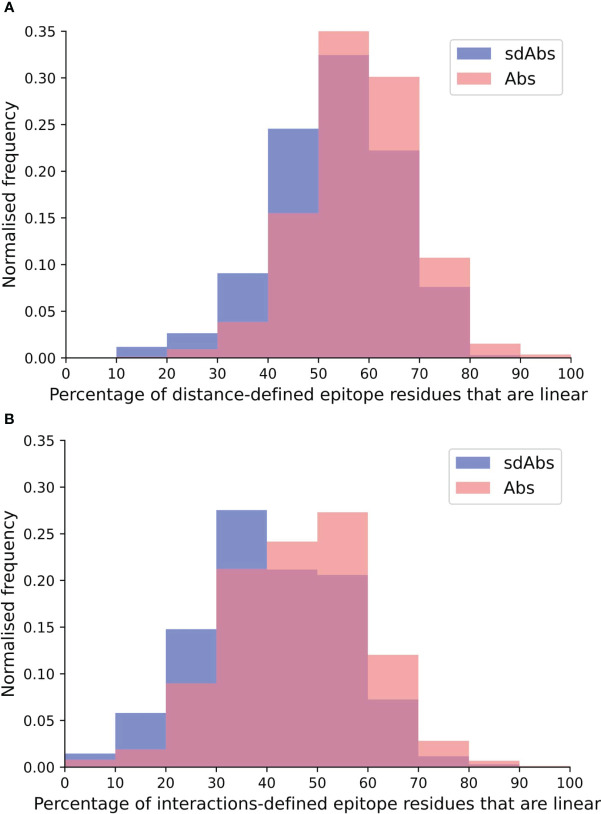
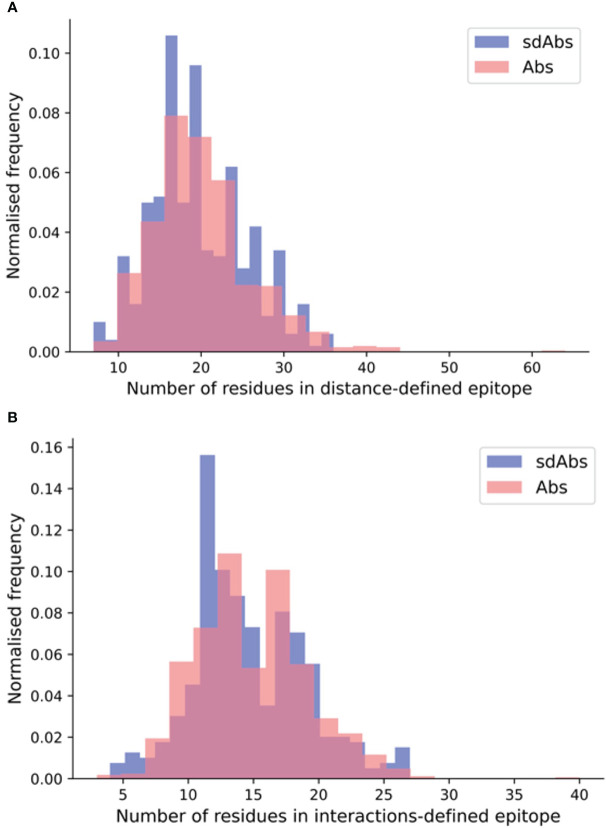
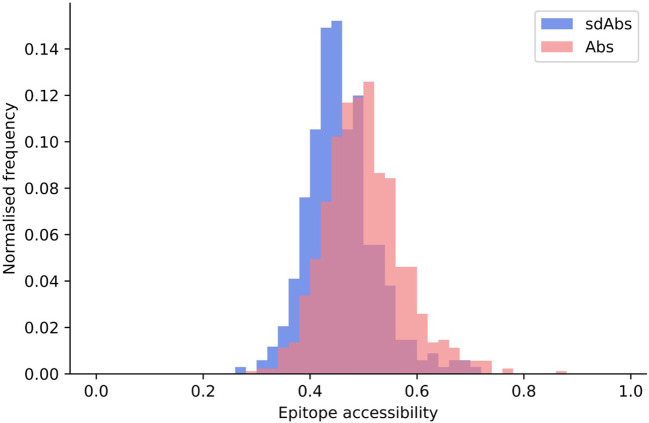
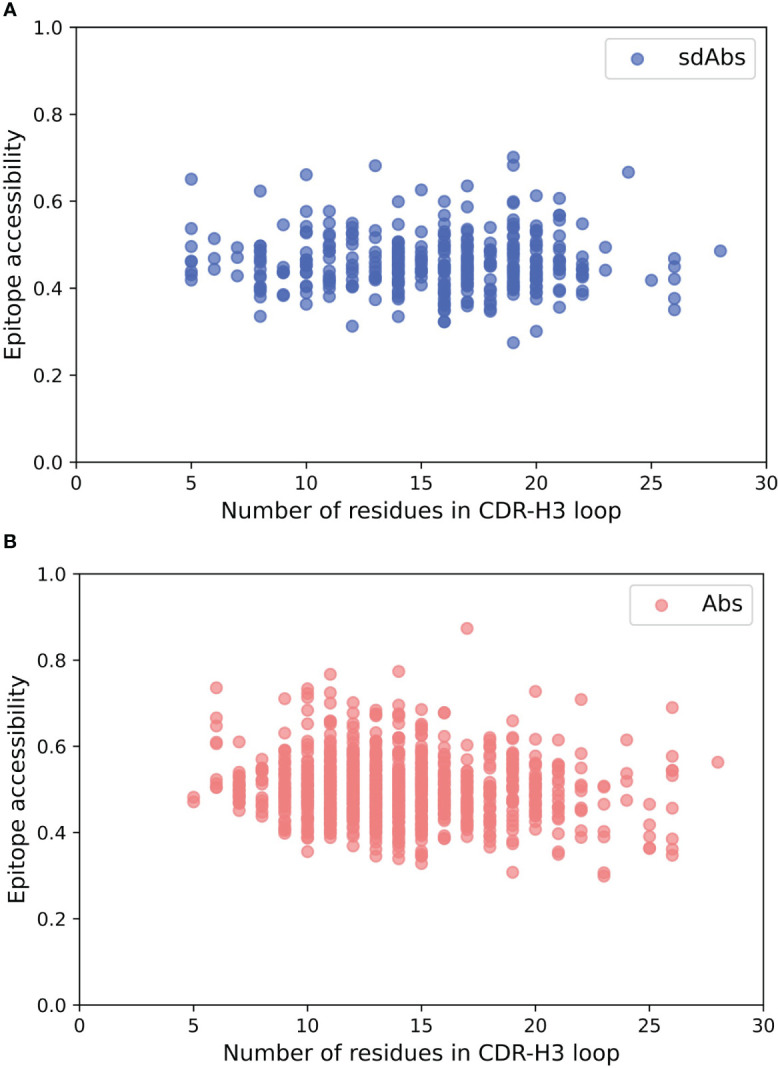
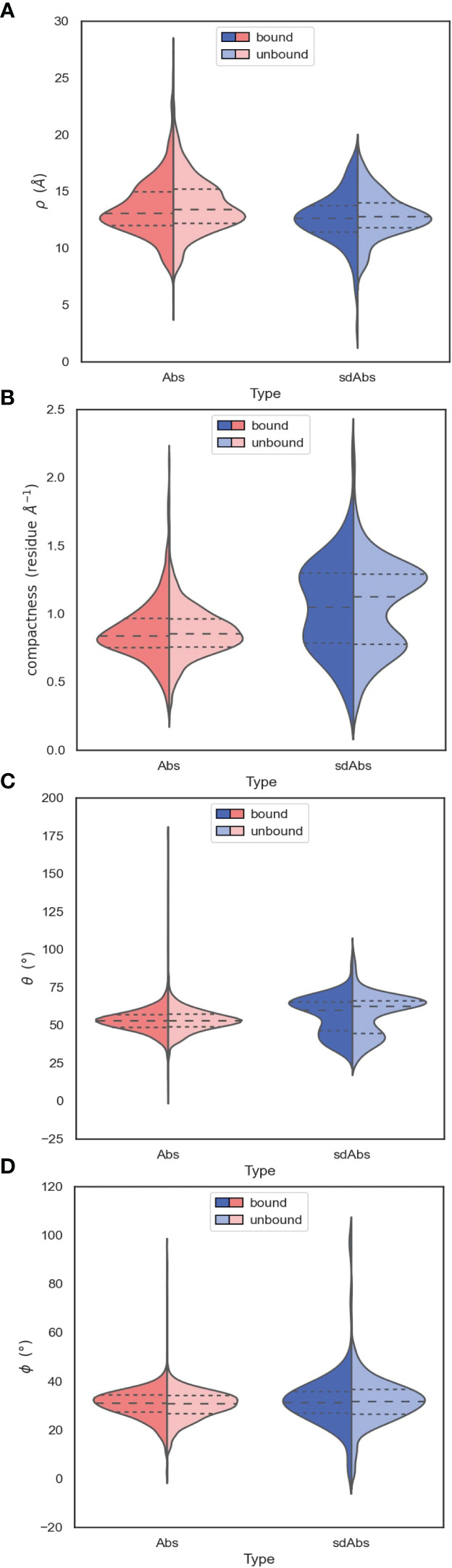
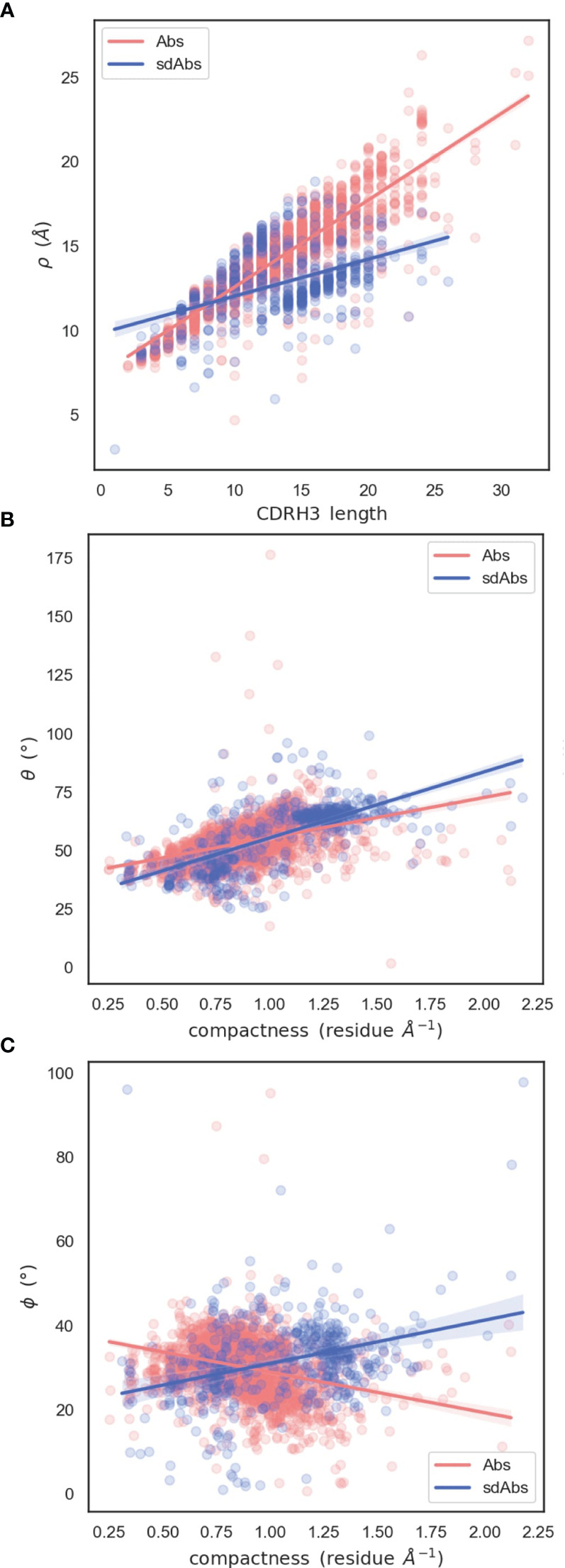
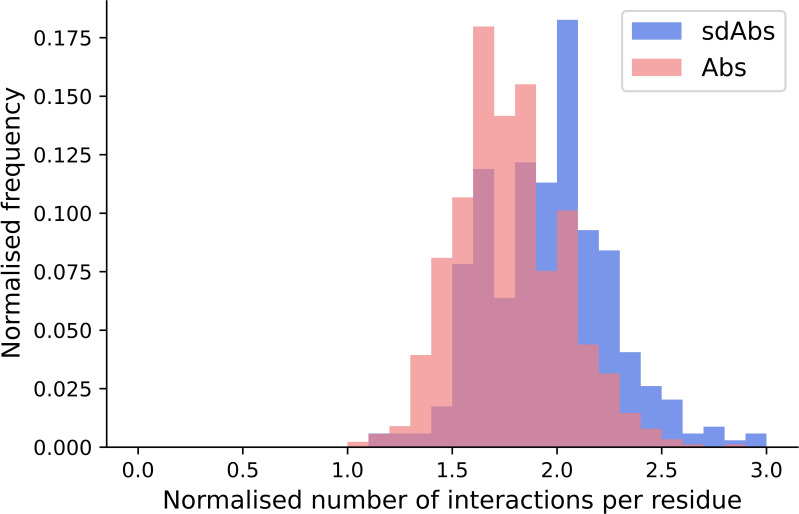
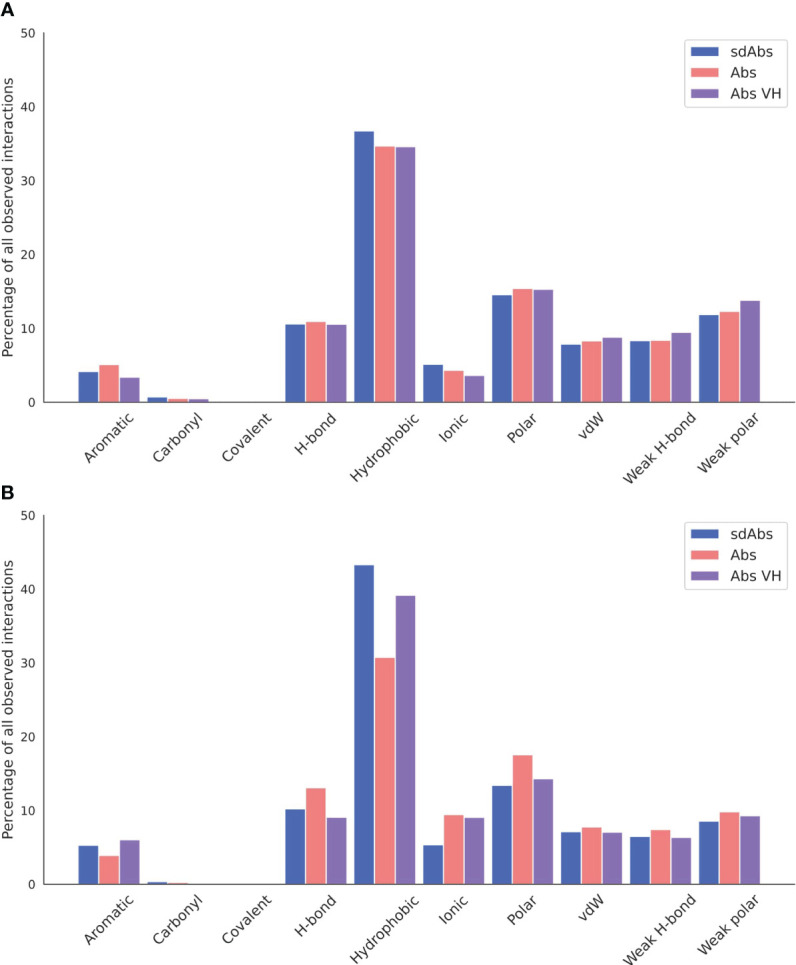
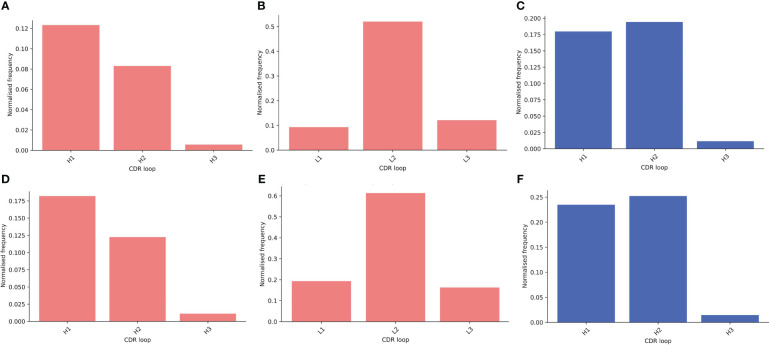
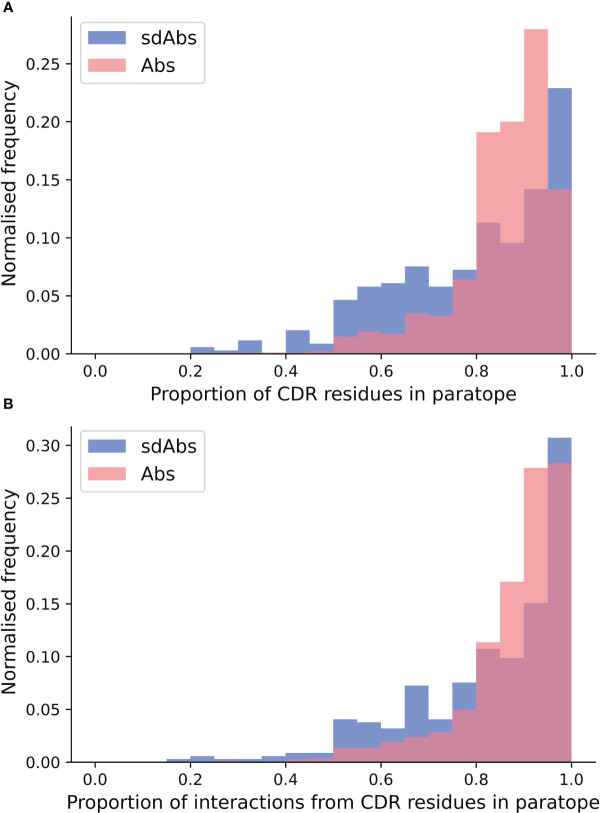
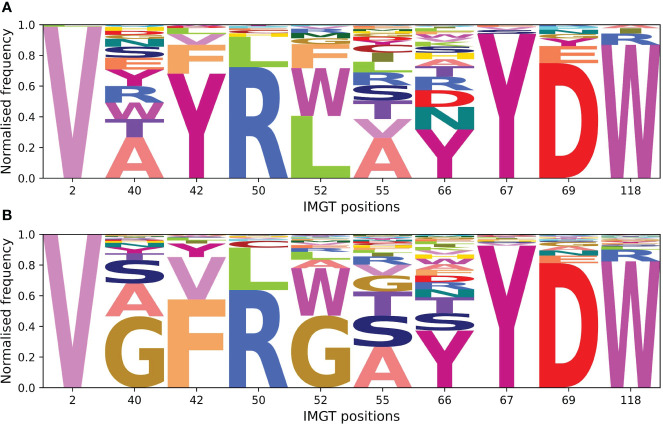
Antibodies are the largest class of biotherapeutics. However, in recent years, single-domain antibodies have gained traction due to their smaller size and comparable binding affinity. Antibodies (Abs) and single-domain antibodies (sdAbs) differ in the structures of their binding sites: most significantly, single-domain antibodies lack a light chain and so have just three CDR loops. Given this inherent structural difference, it is important to understand whether Abs and sdAbs are distinguishable in how they engage a binding partner and thus, whether they are suited to different types of epitopes. In this study, we use non-redundant sequence and structural datasets to compare the paratopes, epitopes and antigen interactions of Abs and sdAbs. We demonstrate that even though sdAbs have smaller paratopes, they target epitopes of equal size to those targeted by Abs. To achieve this, the paratopes of sdAbs contribute more interactions per residue than the paratopes of Abs. Additionally, we find that conserved framework residues are of increased importance in the paratopes of sdAbs, suggesting that they include non-specific interactions to achieve comparable affinity. Furthermore, the epitopes of sdAbs are only marginally less accessible than those of Abs: we posit that this may be explained by differences in the orientation and compaction of sdAb and Ab CDR-H3 loops. Overall, our results have important implications for the engineering and humanization of sdAbs, as well as the selection of the best modality for targeting a particular epitope.
抗体是最大的生物治疗药物类别。然而,近年来,由于单域抗体体积更小且结合亲和力相当,因此受到关注。抗体 (Abs) 和单域抗体 (sdAbs) 在其结合位点的结构上有所不同:最重要的是,单域抗体缺乏轻链,因此只有三个 CDR 环。鉴于这种固有的结构差异,了解 Abs 和 sdAbs 在与结合伴侣结合的方式上是否可区分,以及它们是否适用于不同类型的表位,这一点非常重要。在这项研究中,我们使用非冗余序列和结构数据集来比较 Abs 和 sdAbs 的变区、表位和抗原相互作用。我们证明,尽管 sdAbs 的变区较小,但它们针对的表位与 Abs 针对的表位大小相同。为了实现这一点,sdAbs 的变区每个残基贡献的相互作用比 Abs 的变区多。此外,我们发现保守的框架残基在 sdAbs 的变区中变得更为重要,这表明它们包括非特异性相互作用以实现相当的亲和力。此外,sdAbs 的表位的可及性仅略低于 Abs 的表位:我们假设这可能是由于 sdAb 和 Abs 的 CDR-H3 环的取向和紧凑性的差异所致。总的来说,我们的研究结果对 sdAbs 的工程化和人源化以及针对特定表位选择最佳模式具有重要意义。