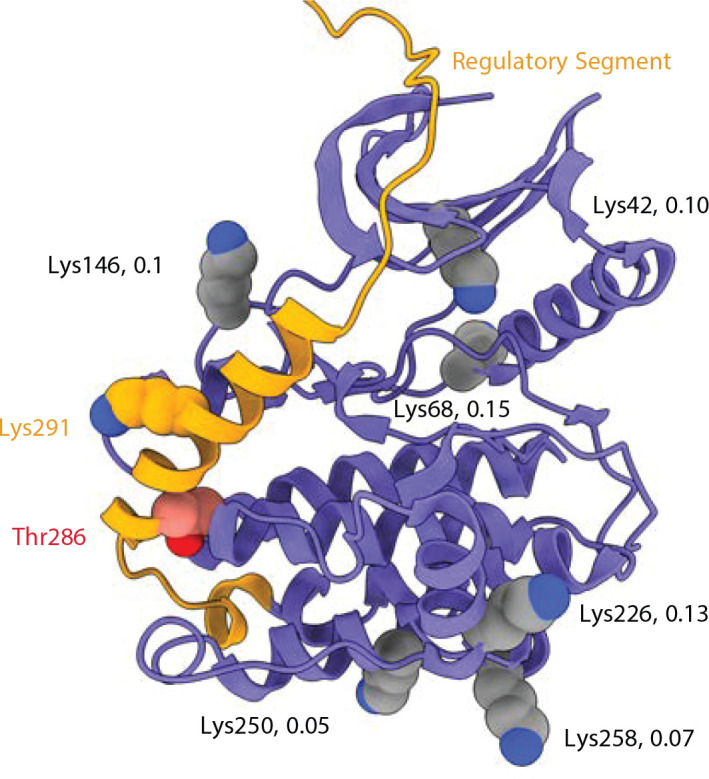
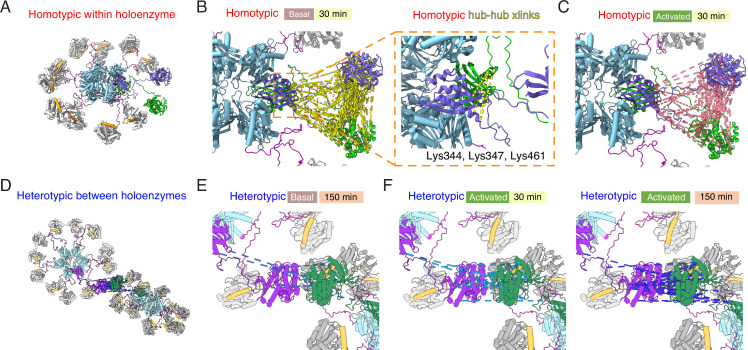
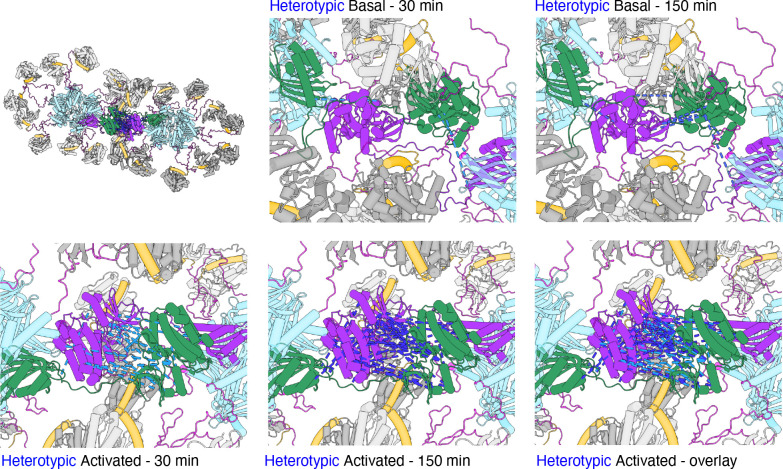
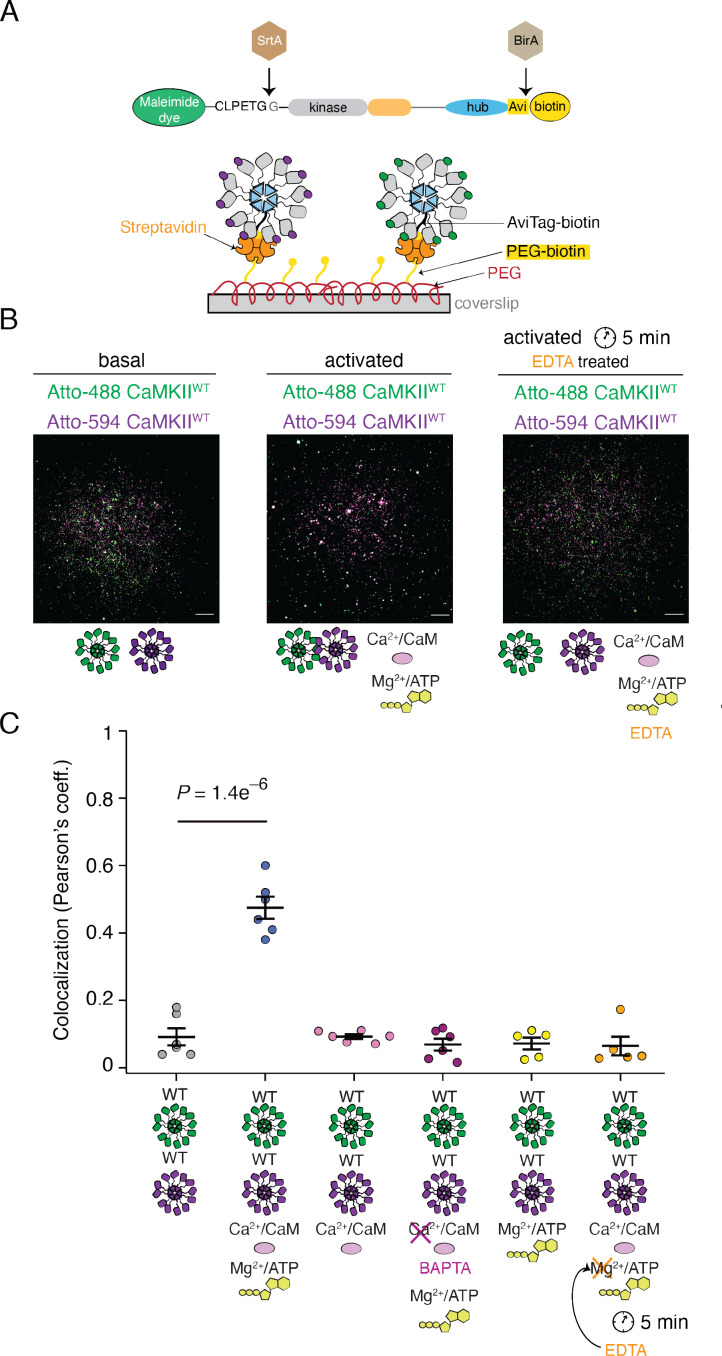
Institute of Biology, Cellular Biophysics, Humboldt Universität zu Berlin, Berlin, Germany.
Leibniz-Forschungsinstitut für Molekulare Pharmakologie, Berlin, Germany.
Elife. 2023 Aug 11;12:e86090. doi: 10.7554/eLife.86090.
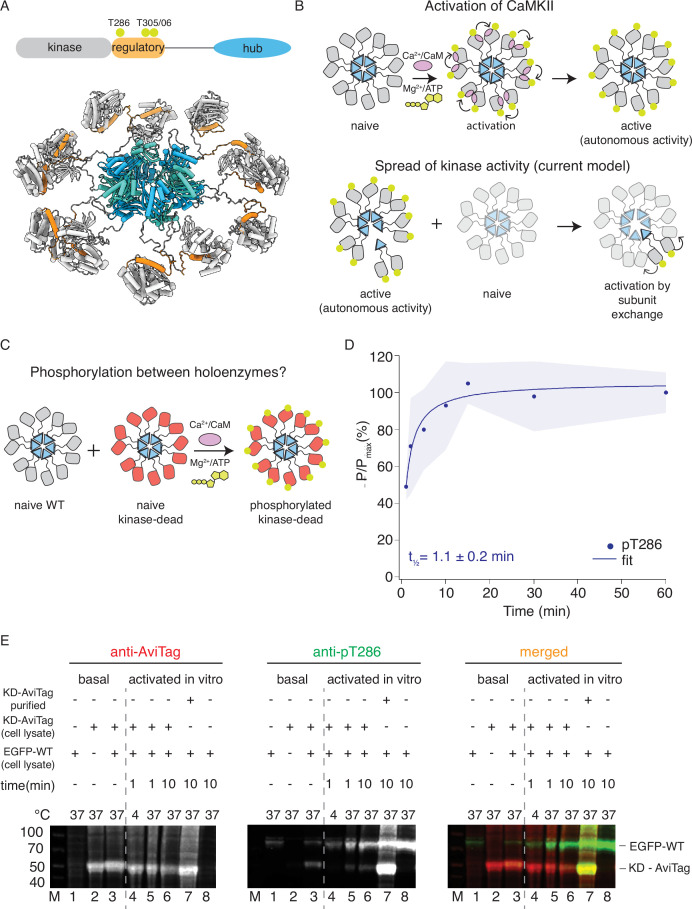
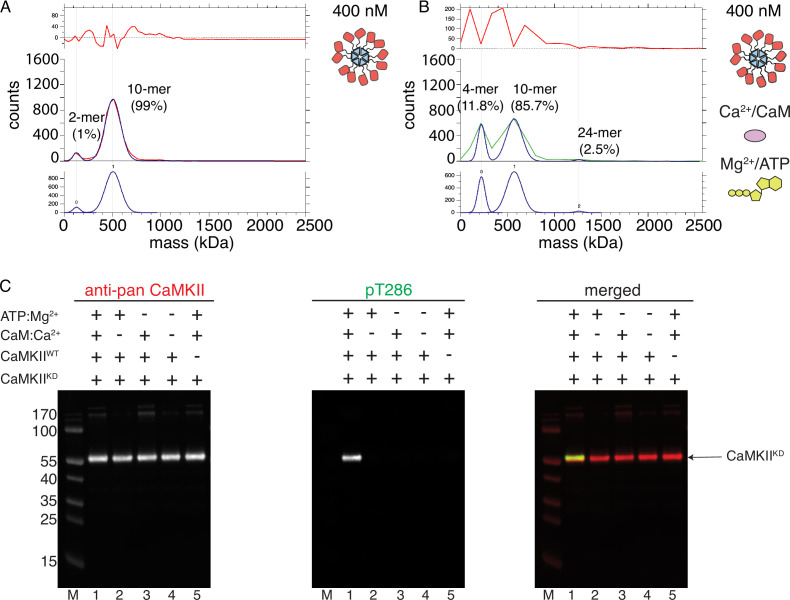
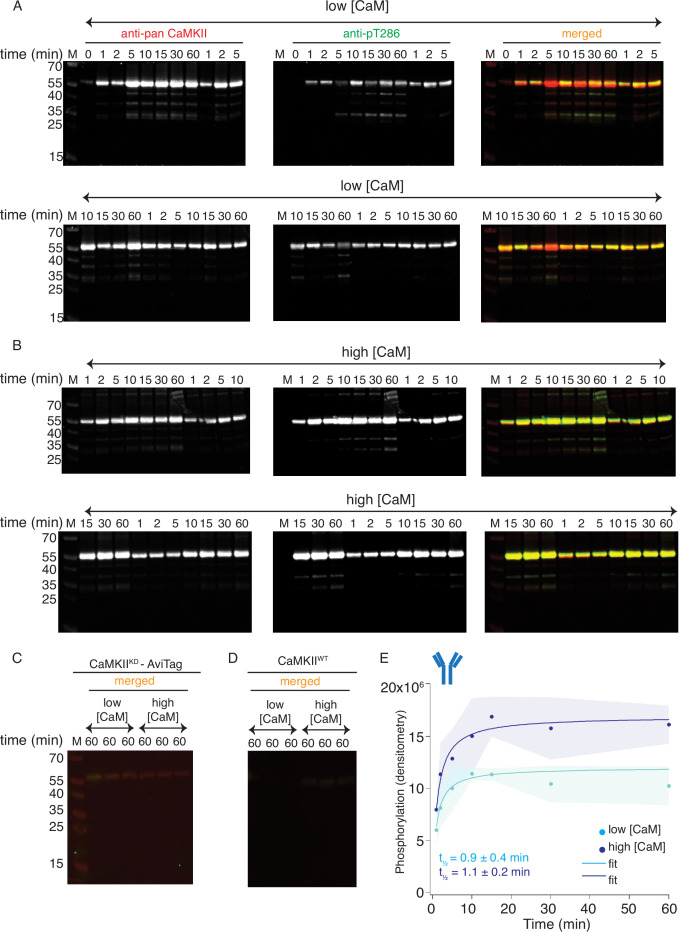
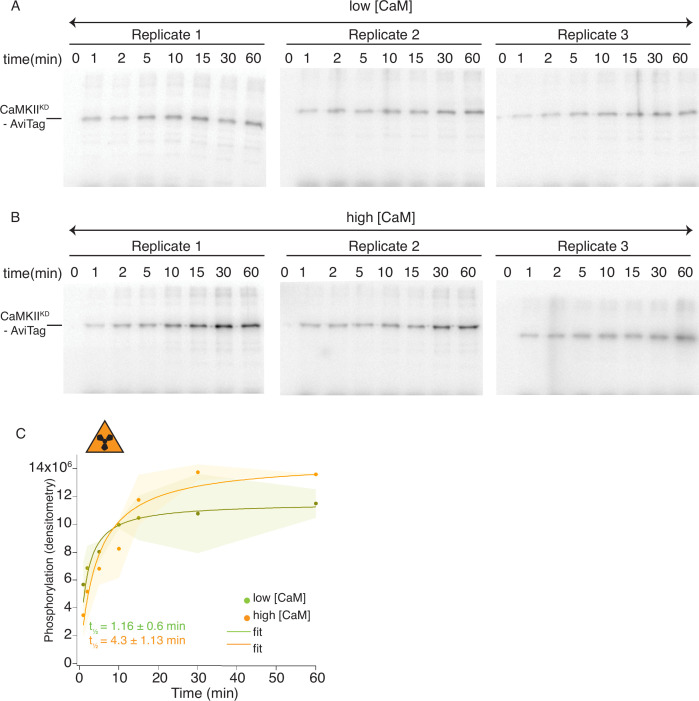
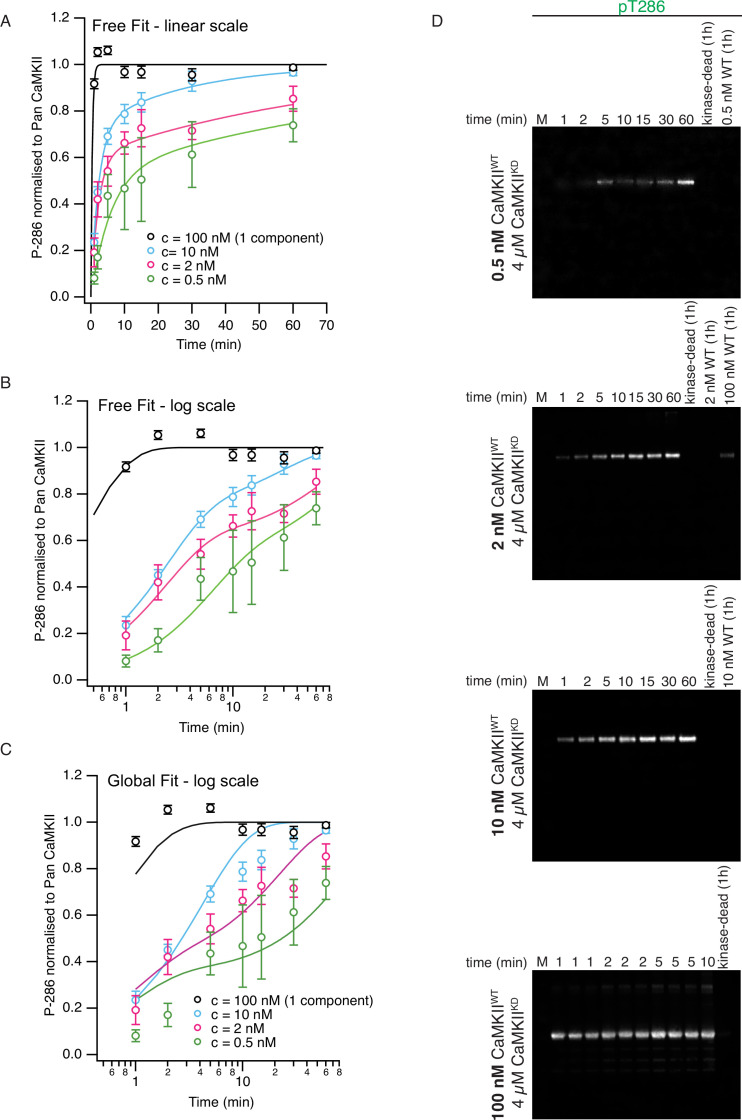
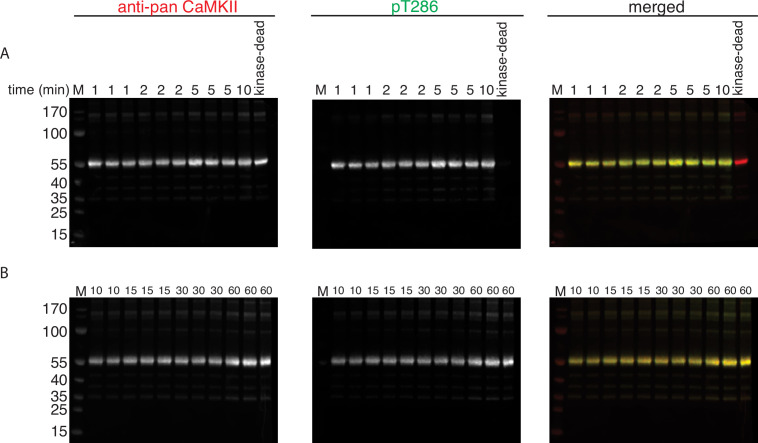
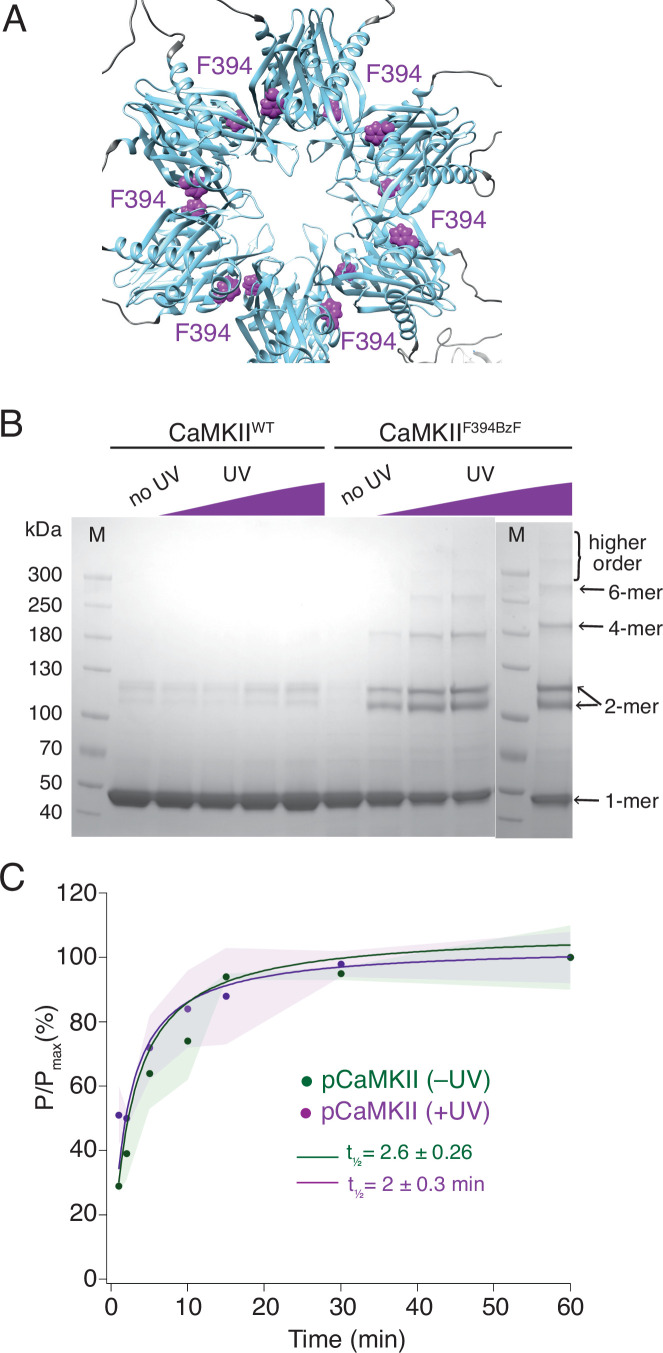
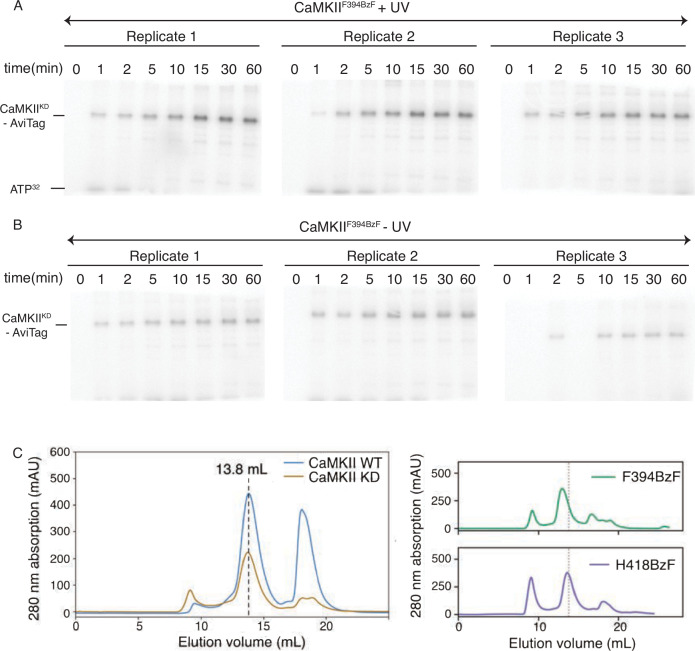
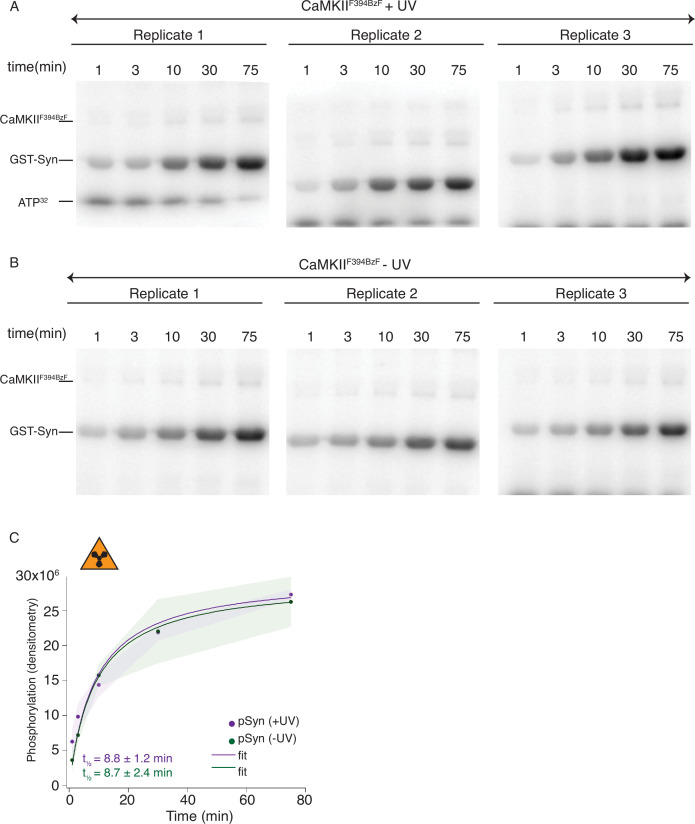
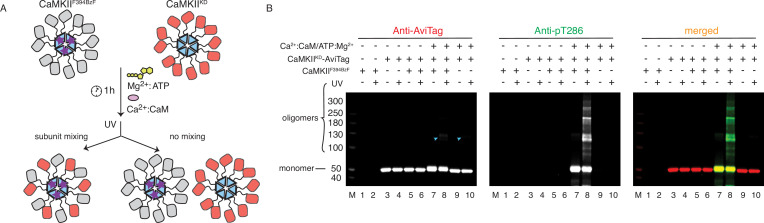
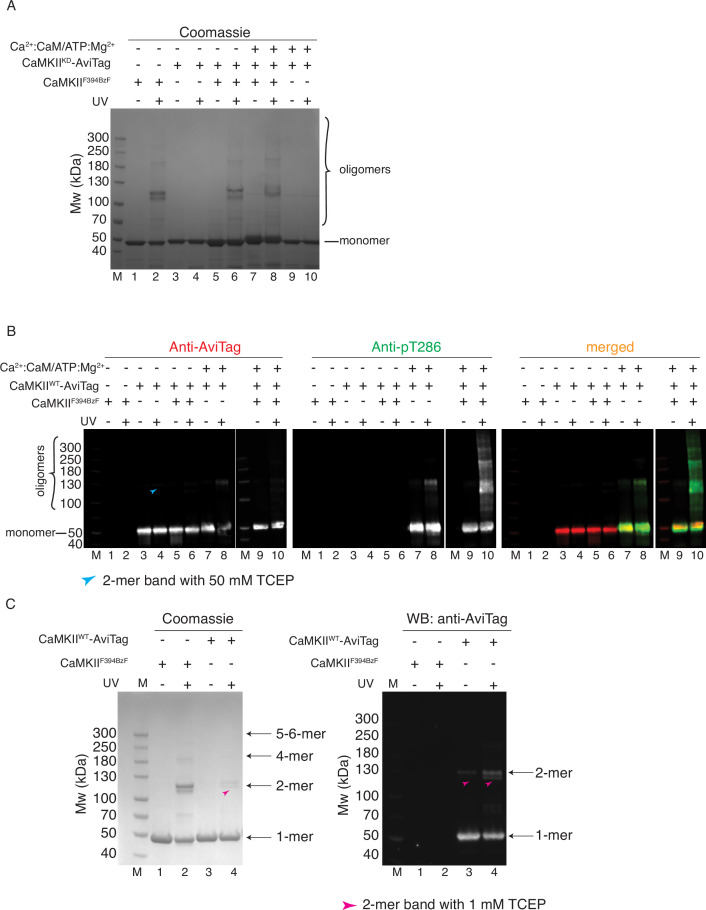
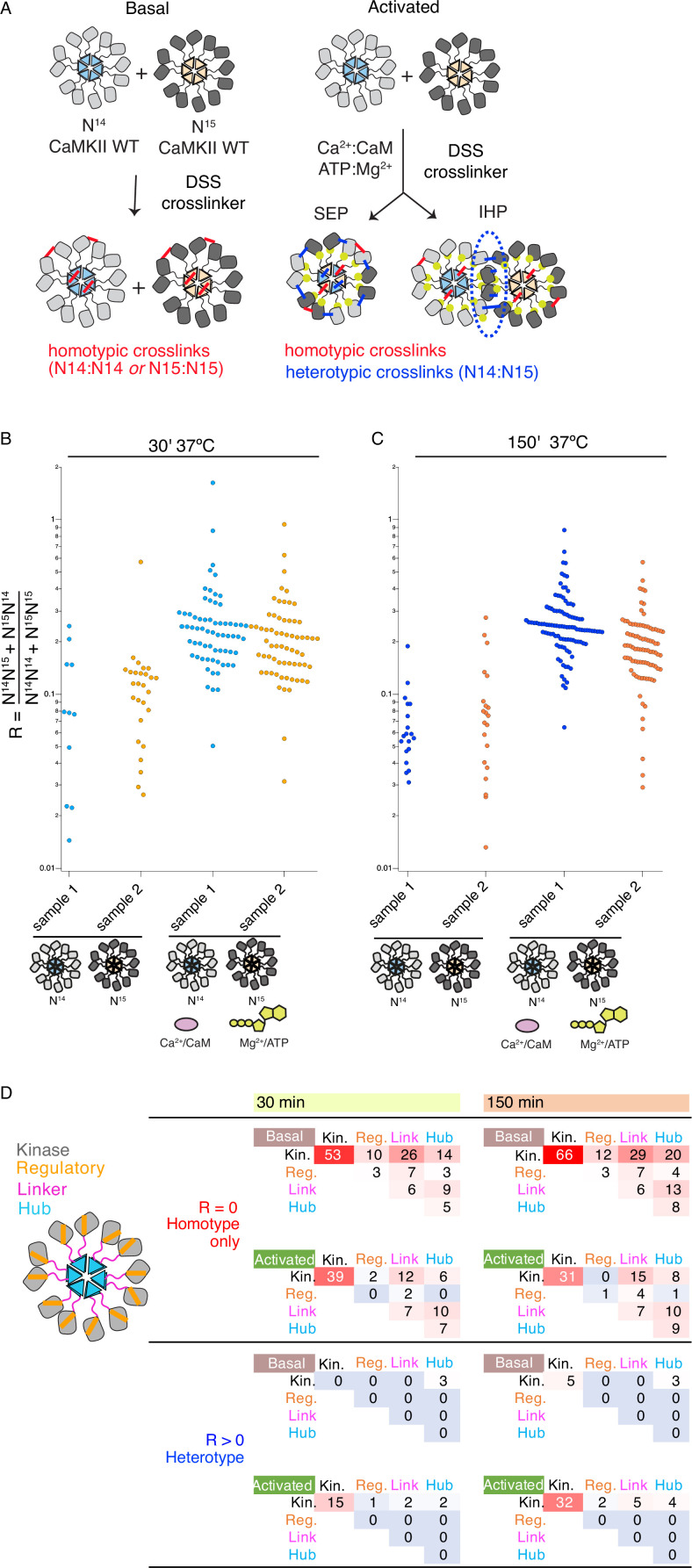
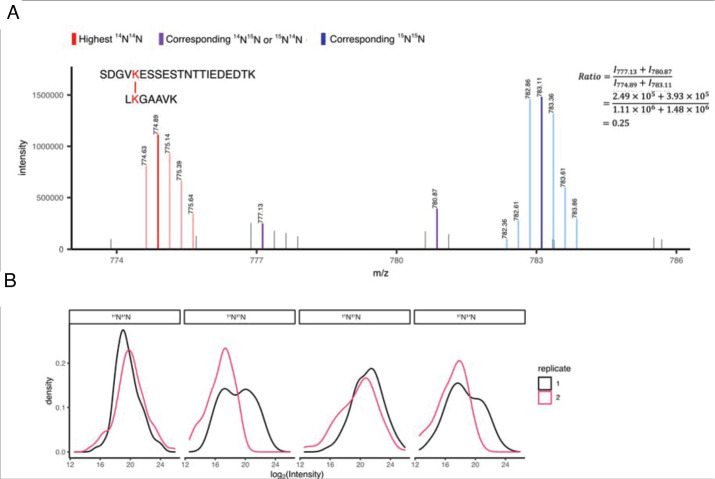
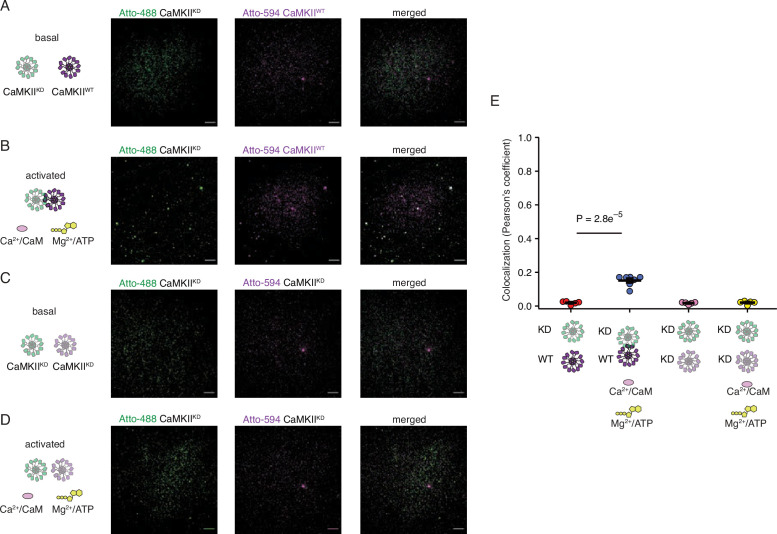
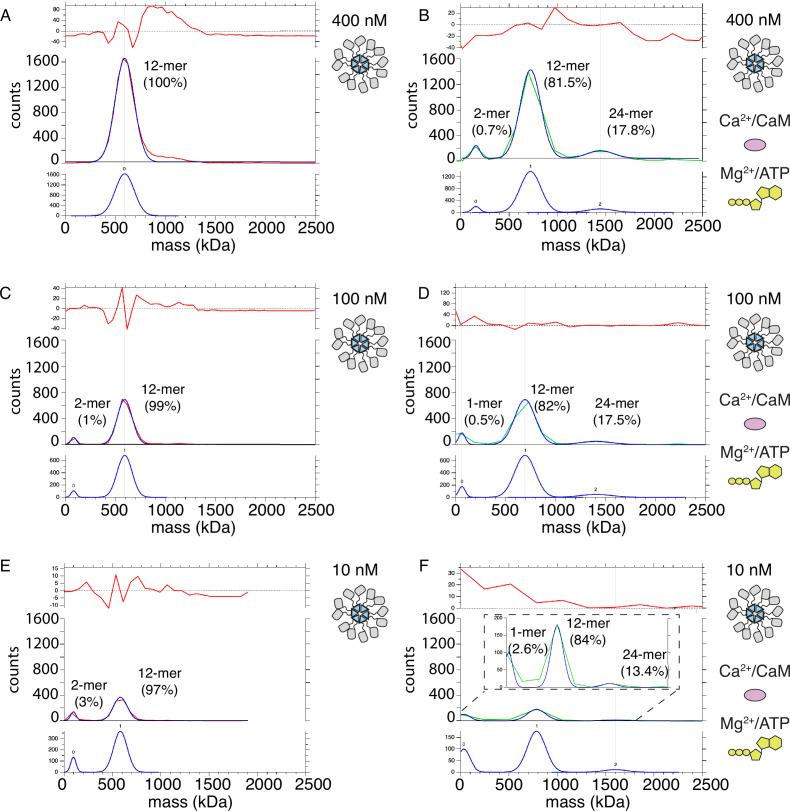
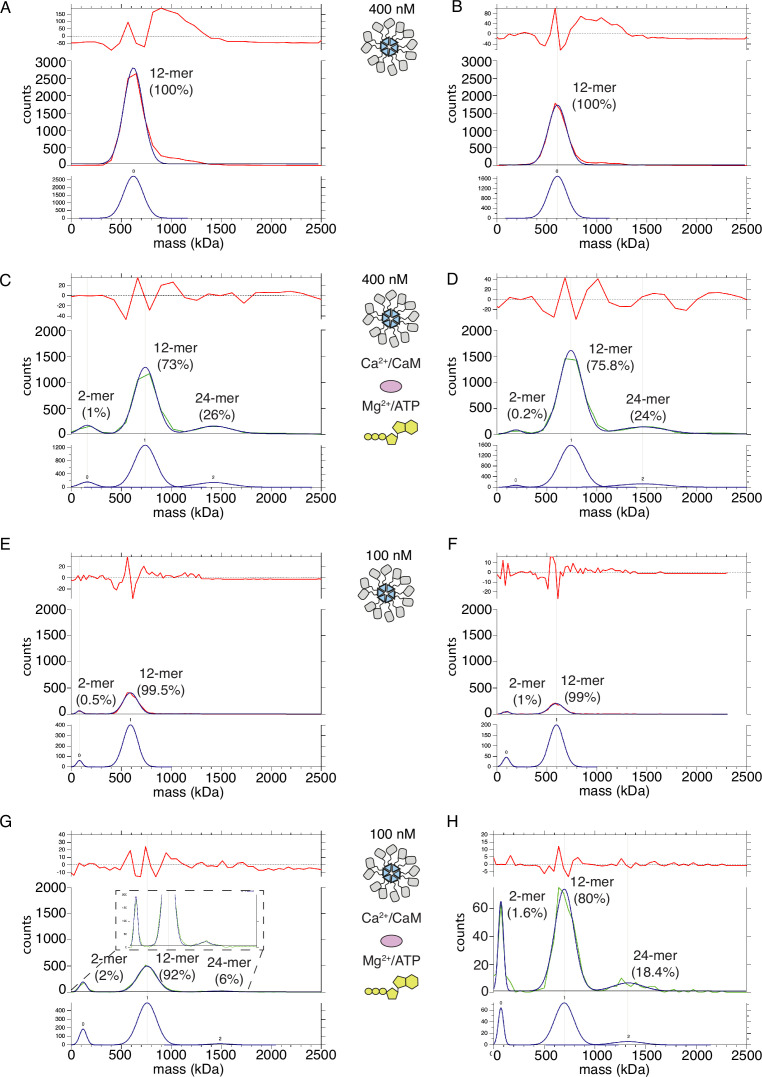
The dodecameric protein kinase CaMKII is expressed throughout the body. The alpha isoform is responsible for synaptic plasticity and participates in memory through its phosphorylation of synaptic proteins. Its elaborate subunit organization and propensity for autophosphorylation allow it to preserve neuronal plasticity across space and time. The prevailing hypothesis for the spread of CaMKII activity, involving shuffling of subunits between activated and naive holoenzymes, is broadly termed subunit exchange. In contrast to the expectations of previous work, we found little evidence for subunit exchange upon activation, and no effect of restraining subunits to their parent holoenzymes. Rather, mass photometry, crosslinking mass spectrometry, single molecule TIRF microscopy and biochemical assays identify inter-holoenzyme phosphorylation (IHP) as the mechanism for spreading phosphorylation. The transient, activity-dependent formation of groups of holoenzymes is well suited to the speed of neuronal activity. Our results place fundamental limits on the activation mechanism of this kinase.
十二聚体蛋白激酶 CaMKII 在体内广泛表达。α 同工型负责突触可塑性,并通过其对突触蛋白的磷酸化参与记忆。其精细的亚基组织和自动磷酸化倾向使其能够在空间和时间上保持神经元可塑性。涉及激活和幼稚全酶之间亚基 shuffling 的 CaMKII 活性传播的流行假说通常被称为亚基交换。与之前工作的预期相反,我们在激活时几乎没有发现亚基交换的证据,也没有将亚基限制在其亲本全酶中的效果。相反,质量光度法、交联质谱法、单分子 TIRF 显微镜和生化测定将同种酶间磷酸化(IHP)鉴定为磷酸化传播的机制。全酶组的短暂、活性依赖性形成非常适合神经元活动的速度。我们的结果对这种激酶的激活机制施加了基本限制。