Computational Biology Section, Laboratory of Immune System Biology, National Institute of Allergy and Infectious Diseases, National Institutes of Health, Bethesda, United States.
Elife. 2023 Oct 20;12:e90681. doi: 10.7554/eLife.90681.
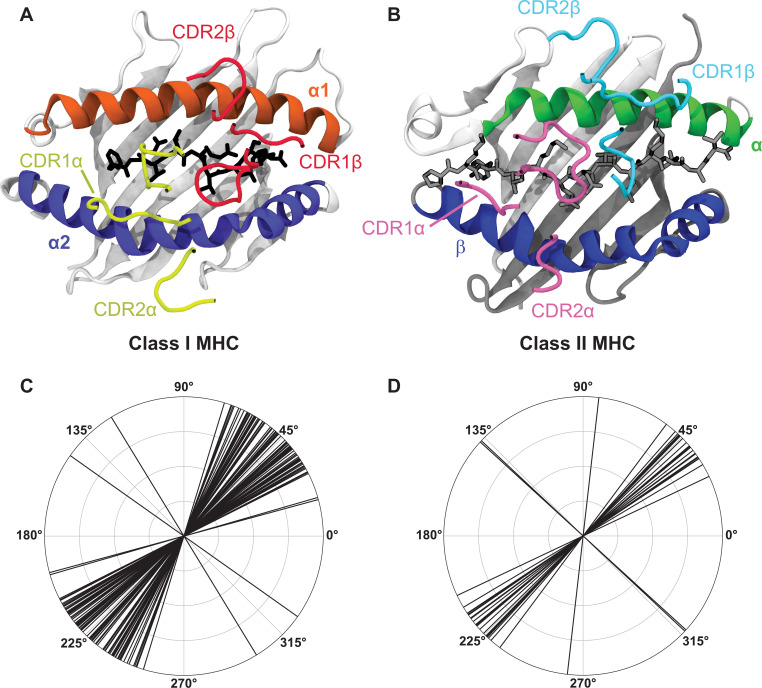
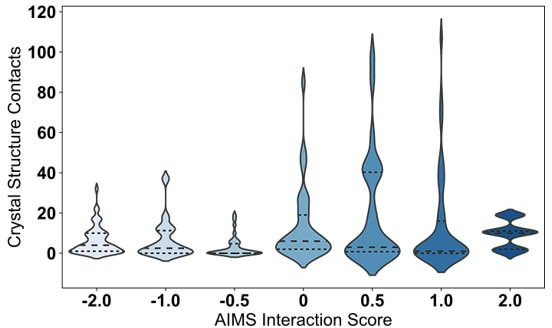
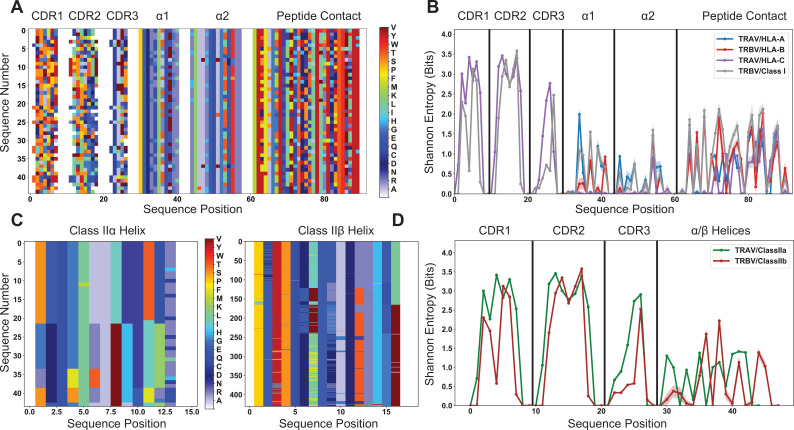
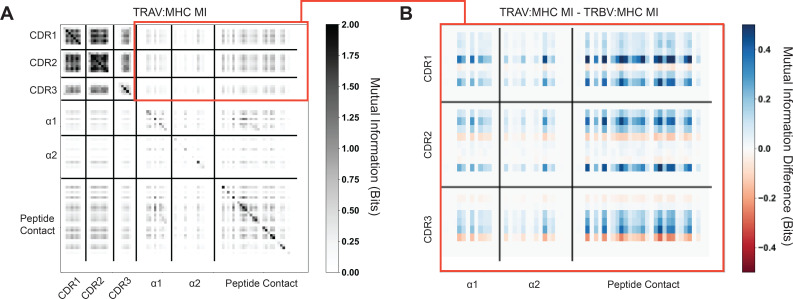
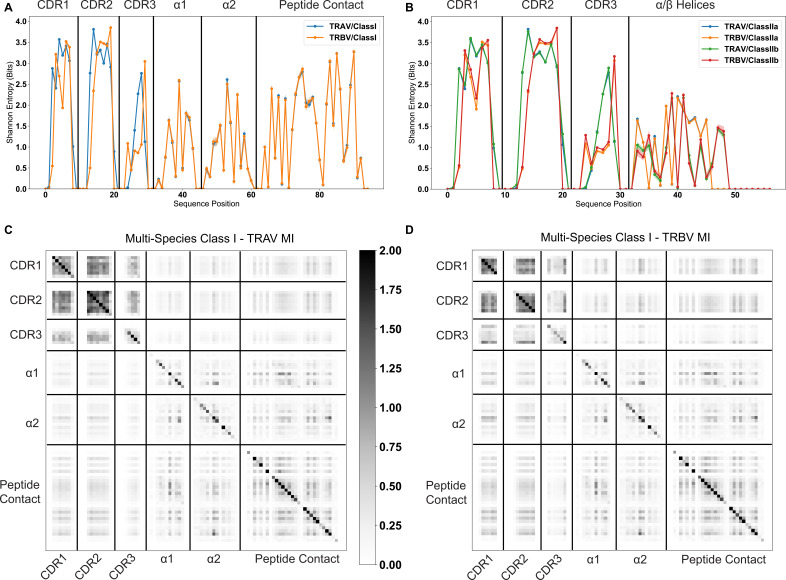
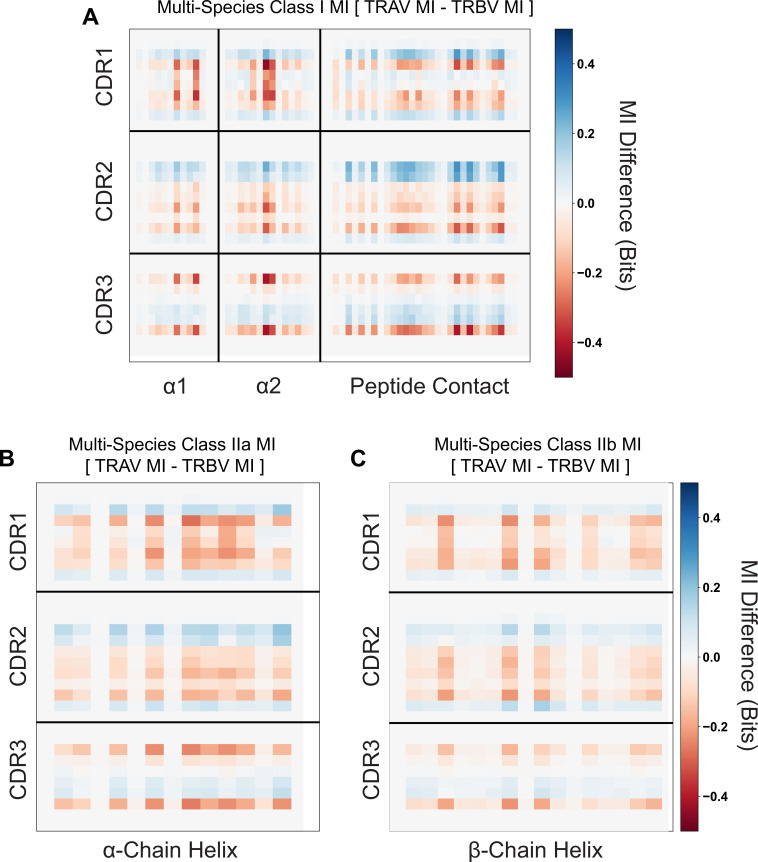
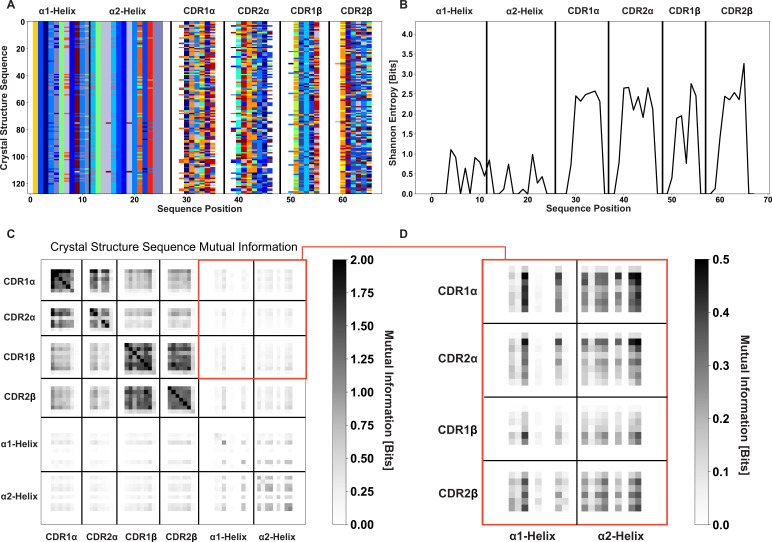
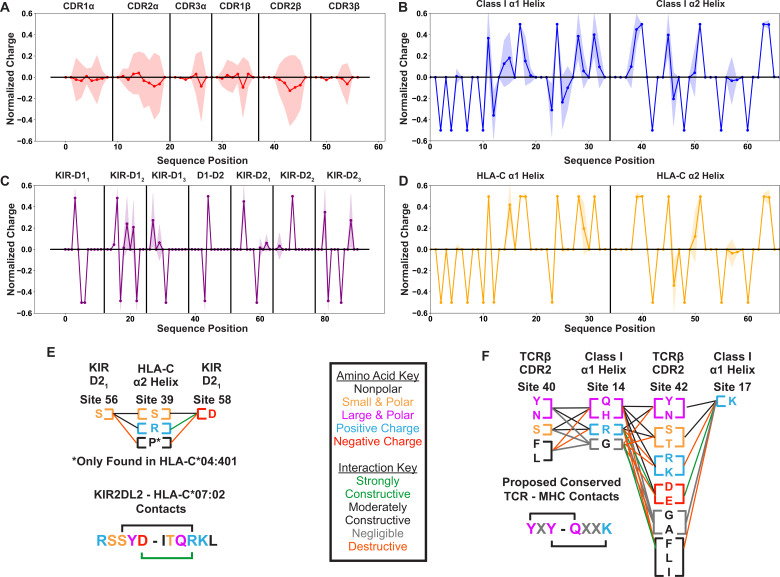
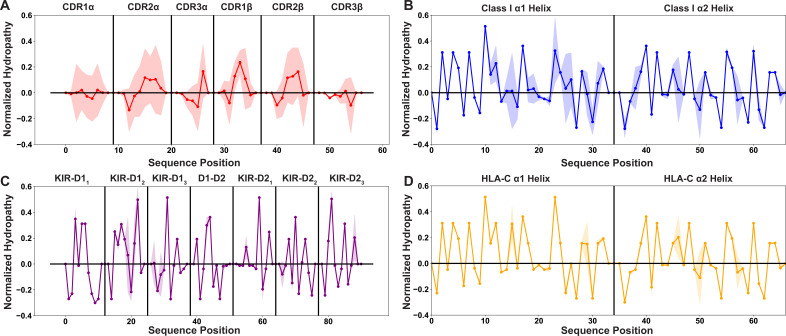
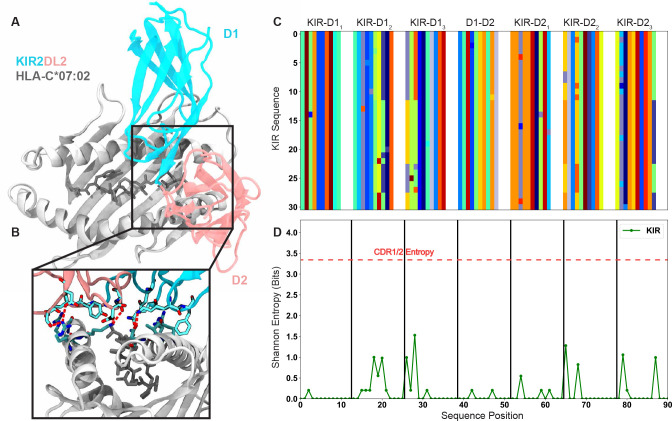
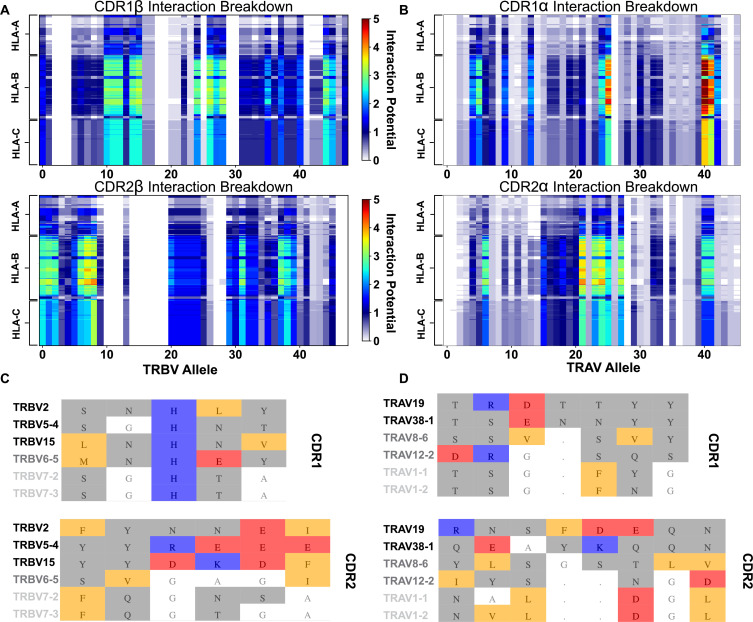
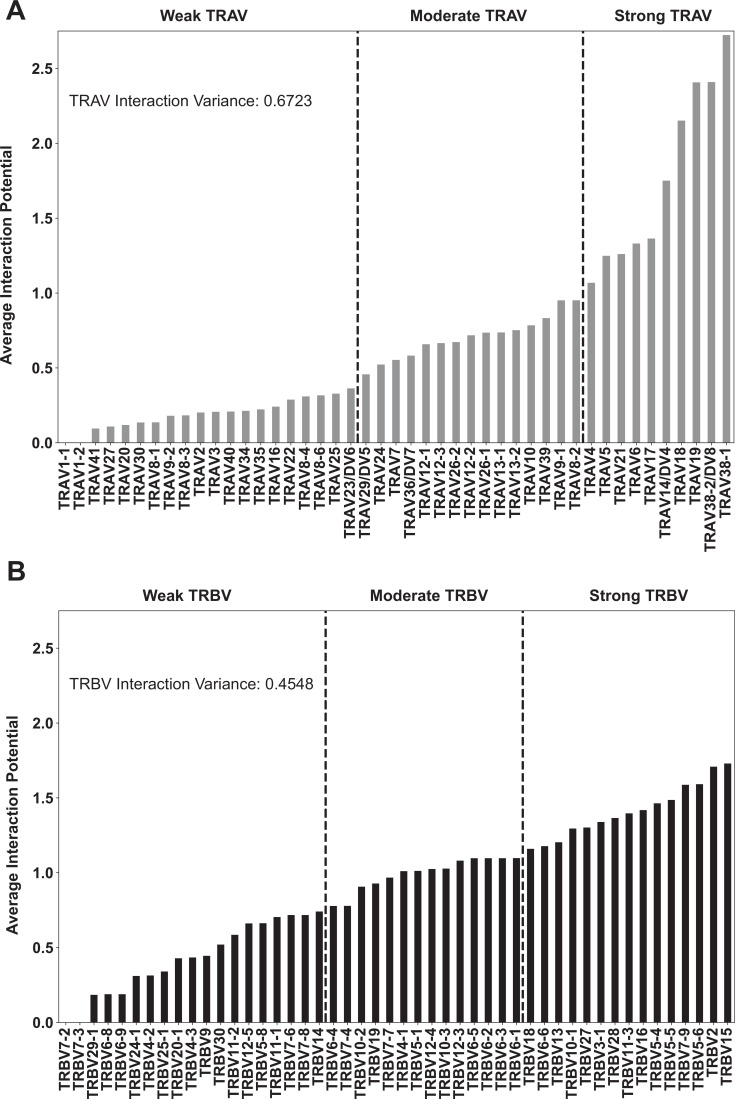
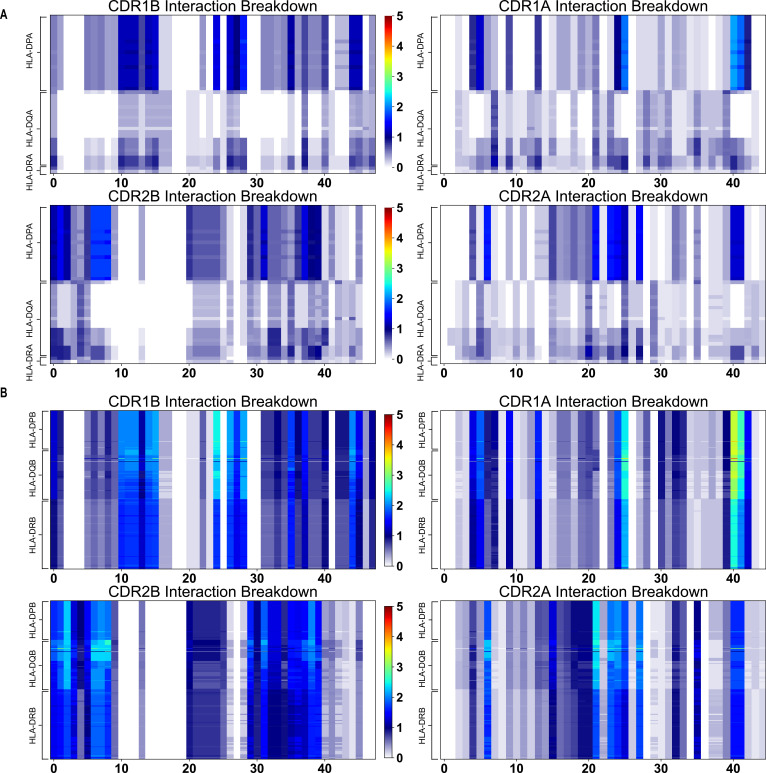
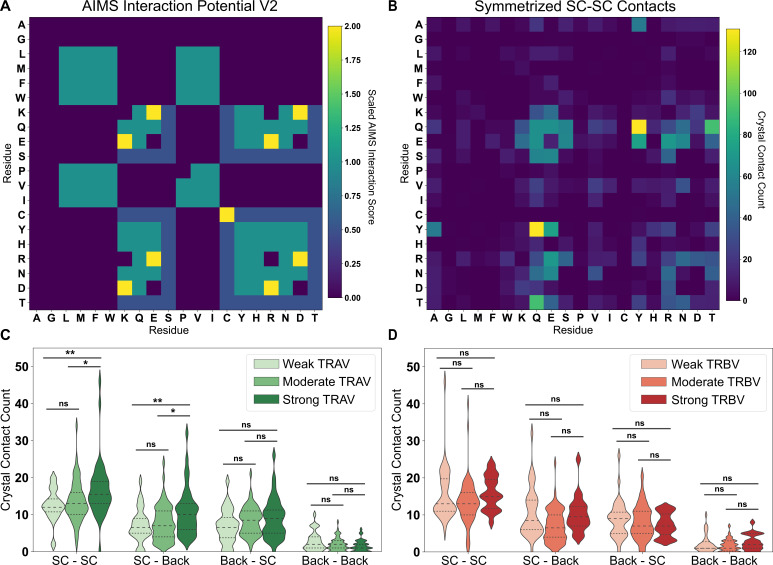
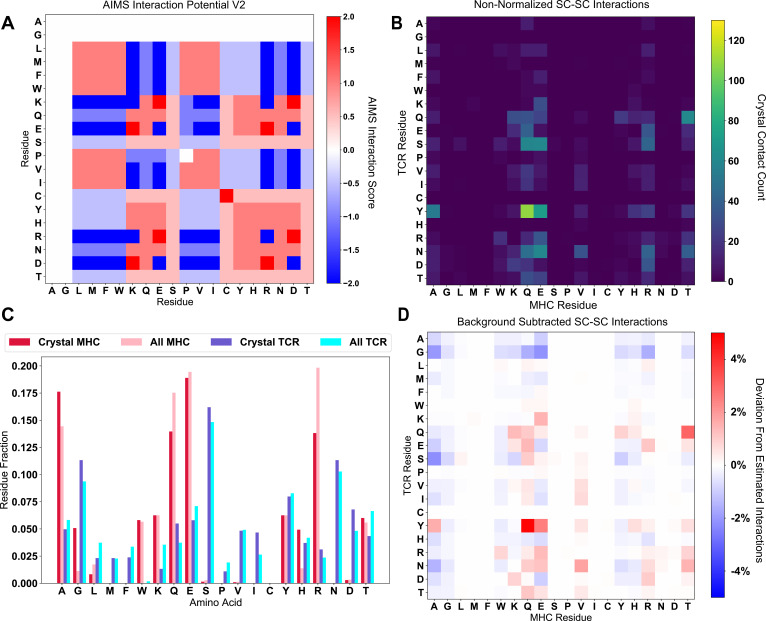
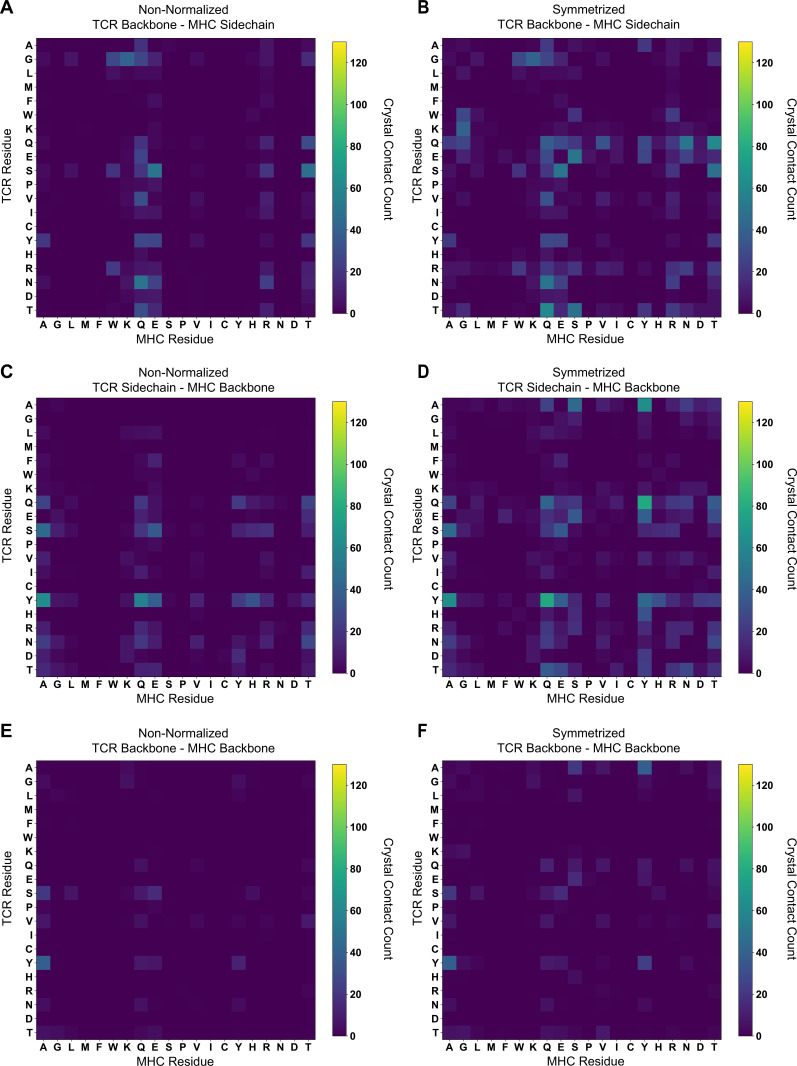
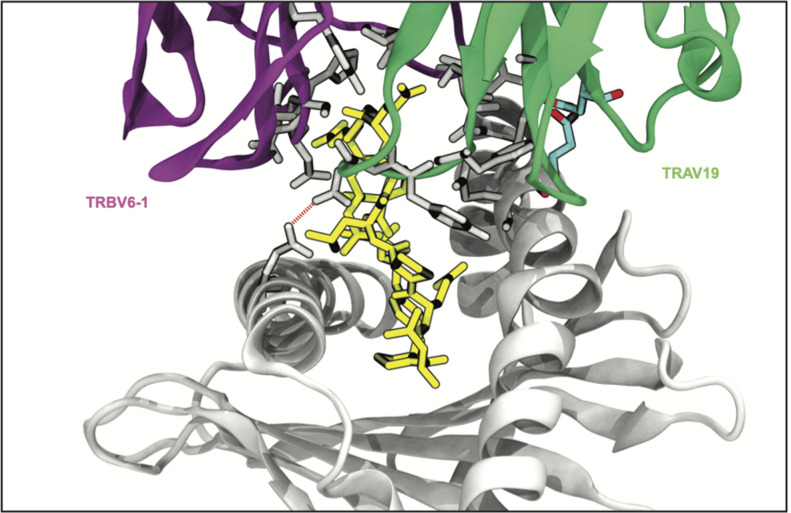
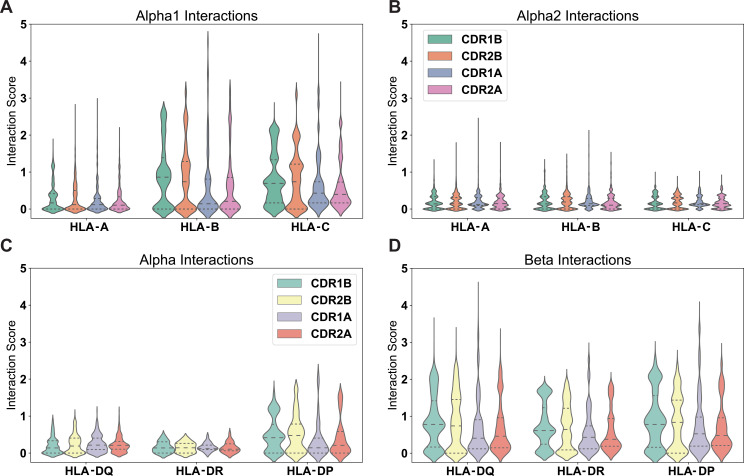
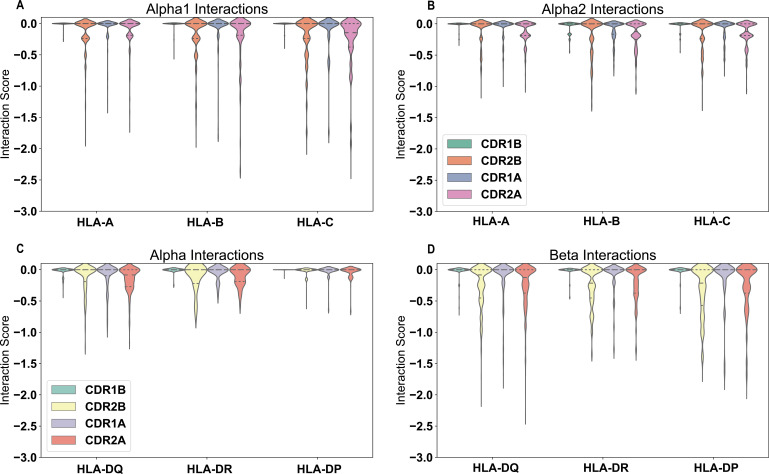
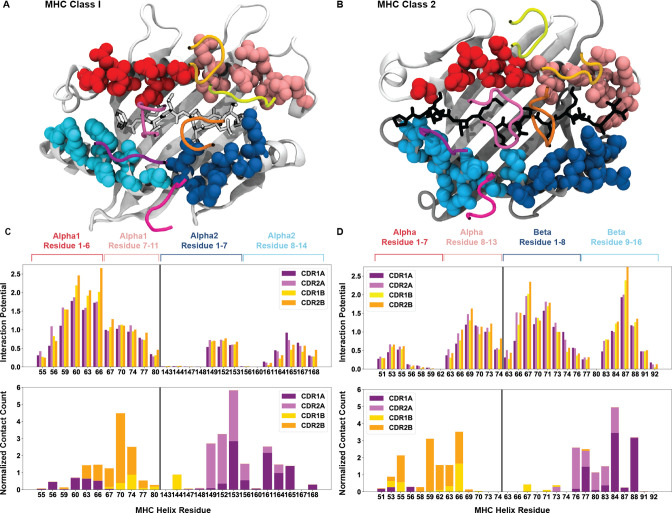
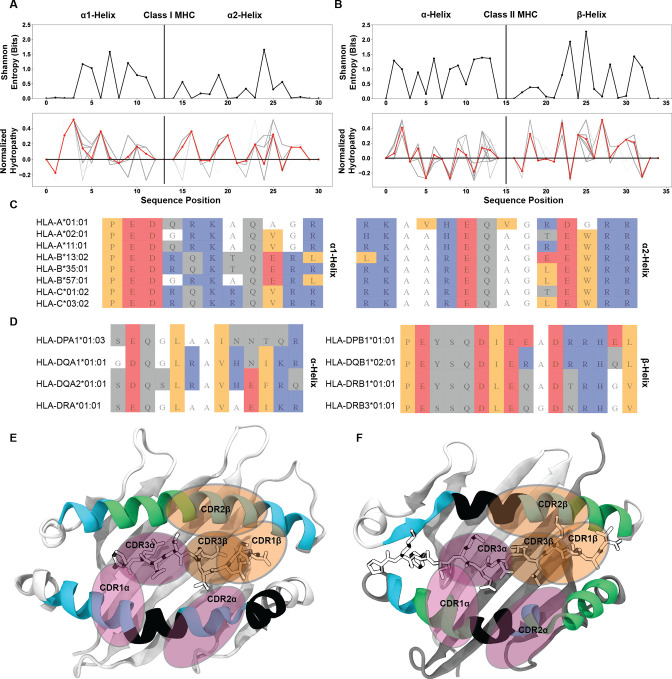
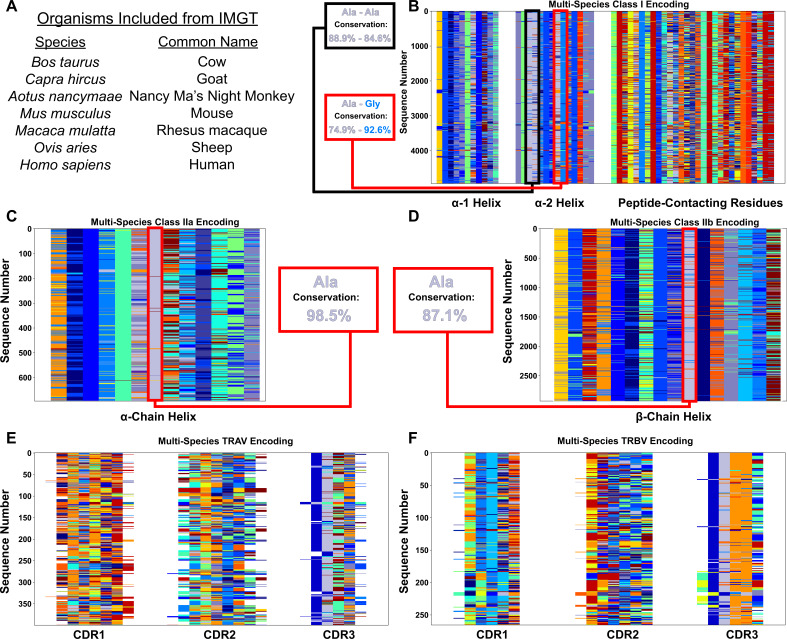
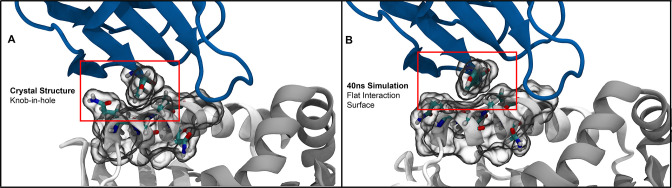
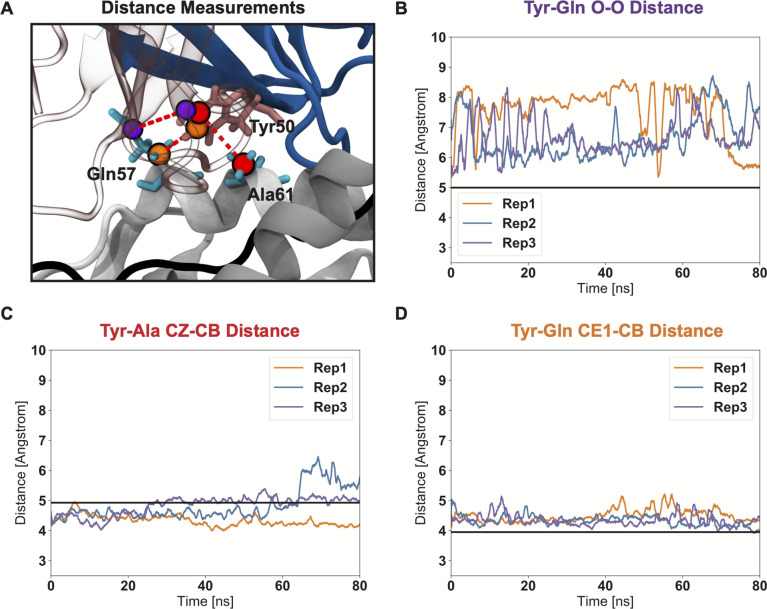
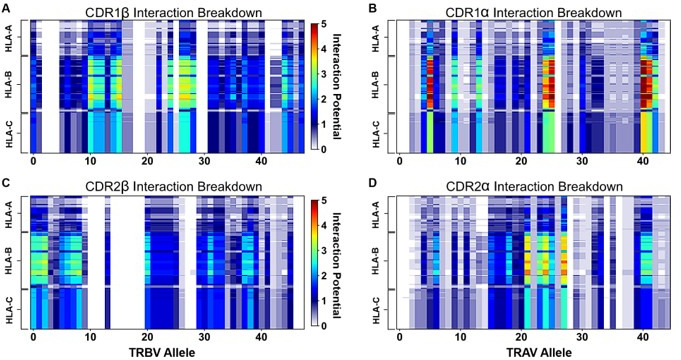
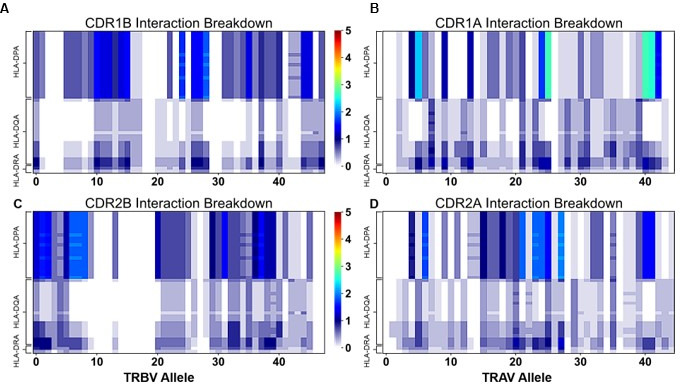
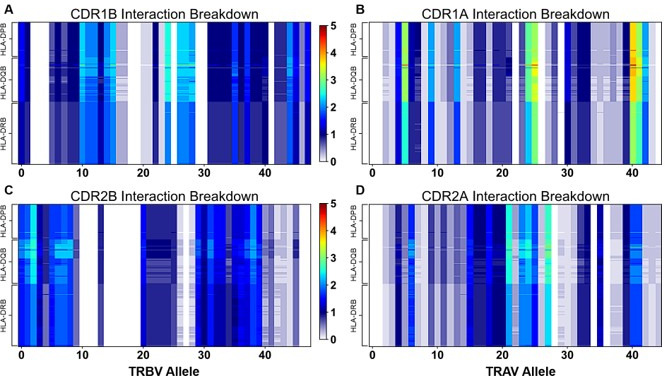
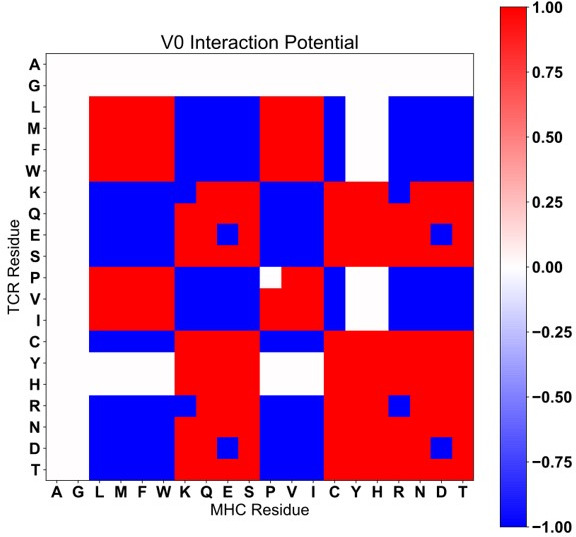
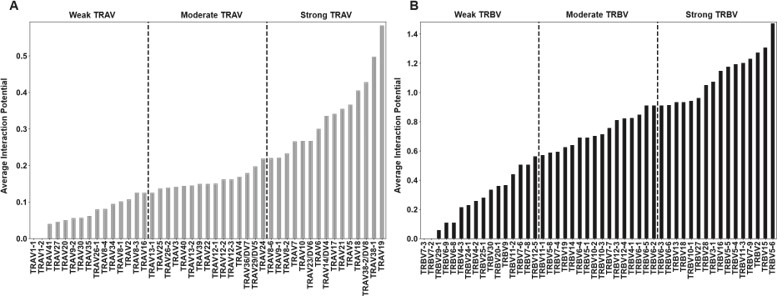
T cells are critically important components of the adaptive immune system primarily responsible for identifying and responding to pathogenic challenges. This recognition of pathogens is driven by the interaction between membrane-bound T cell receptors (TCRs) and antigenic peptides presented on major histocompatibility complex (MHC) molecules. The formation of the TCR-peptide-MHC complex (TCR-pMHC) involves interactions among germline-encoded and hypervariable amino acids. Germline-encoded and hypervariable regions can form contacts critical for complex formation, but only interactions between germline-encoded contacts are likely to be shared across many of all the possible productive TCR-pMHC complexes. Despite this, experimental investigation of these interactions have focused on only a small fraction of the possible interaction space. To address this, we analyzed every possible germline-encoded TCR-MHC contact in humans, thereby generating the first comprehensive characterization of these largely antigen-independent interactions. Our computational analysis suggests that germline-encoded TCR-MHC interactions that are conserved at the sequence level are rare due to the high amino acid diversity of the TCR CDR1 and CDR2 loops, and that such conservation is unlikely to dominate the dynamic protein-protein binding interface. Instead, we propose that binding properties such as the docking orientation are defined by regions of biophysical compatibility between these loops and the MHC surface.
T 细胞是适应性免疫系统的重要组成部分,主要负责识别和应对病原体的挑战。这种对病原体的识别是由膜结合的 T 细胞受体(TCR)与主要组织相容性复合体(MHC)分子上呈递的抗原肽之间的相互作用驱动的。TCR-肽-MHC 复合物(TCR-pMHC)的形成涉及种系编码和高变区氨基酸之间的相互作用。种系编码和高变区可以形成对复合物形成至关重要的接触,但只有种系编码接触之间的相互作用才可能在许多可能的有生产力的 TCR-pMHC 复合物中共享。尽管如此,这些相互作用的实验研究仅集中在可能的相互作用空间的一小部分。为了解决这个问题,我们分析了人类中每一个可能的种系编码的 TCR-MHC 接触,从而首次全面描述了这些在很大程度上与抗原无关的相互作用。我们的计算分析表明,由于 TCR CDR1 和 CDR2 环的氨基酸多样性很高,种系编码的 TCR-MHC 相互作用在序列水平上的保守性很少,而且这种保守性不太可能主导动态的蛋白质-蛋白质结合界面。相反,我们提出,结合特性,如对接方向,是由这些环与 MHC 表面之间的物理兼容性区域定义的。