Peter MacCallum Cancer Centre, Melbourne, Vic, Australia.
Sir Peter MacCallum Department of Oncology, The University of Melbourne, Melbourne, Vic, Australia.
EMBO Rep. 2023 Dec 6;24(12):e57695. doi: 10.15252/embr.202357695. Epub 2023 Nov 28.
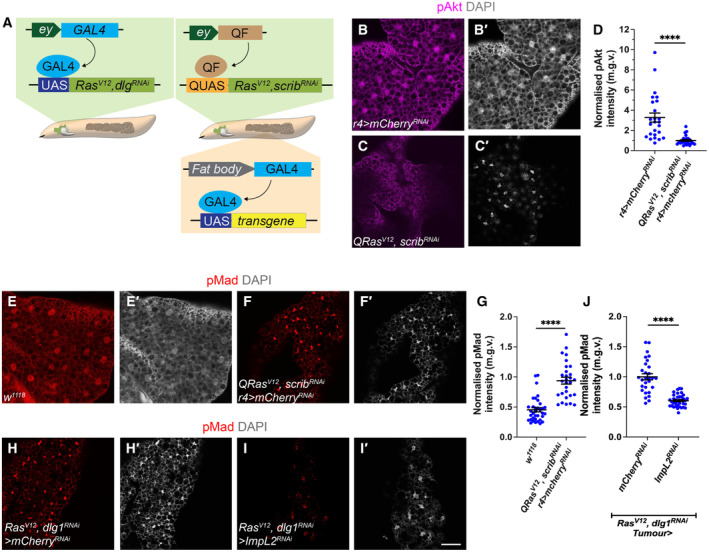
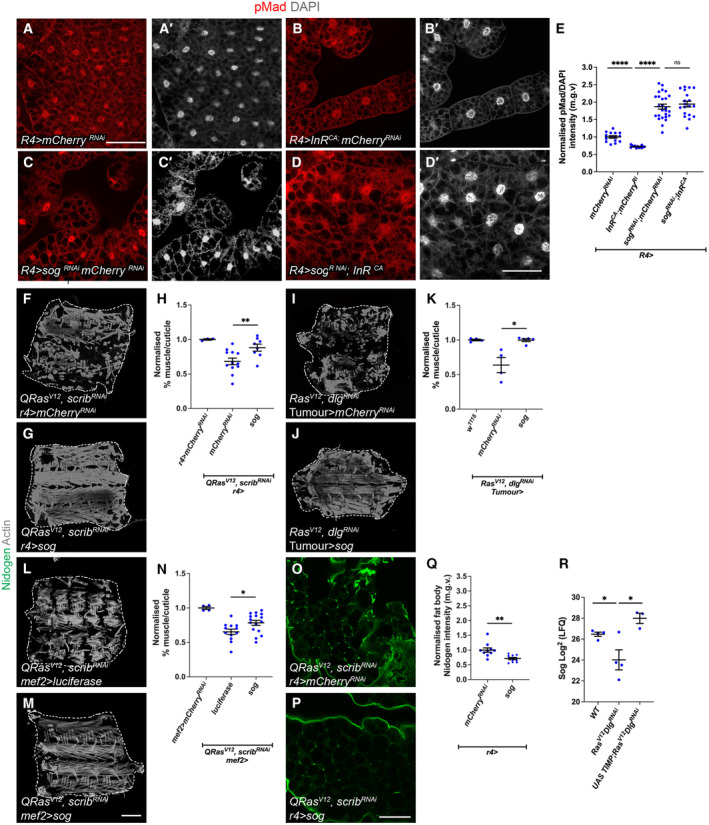
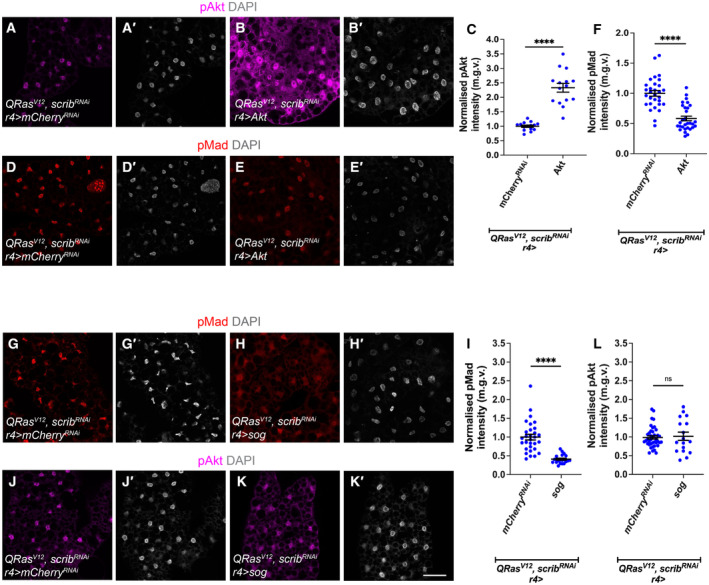
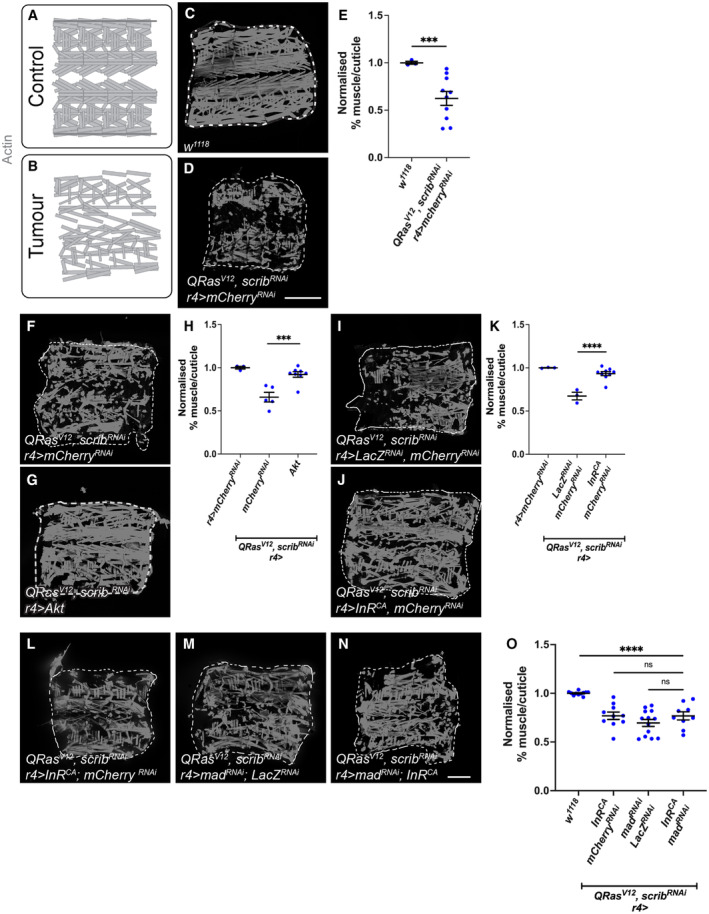
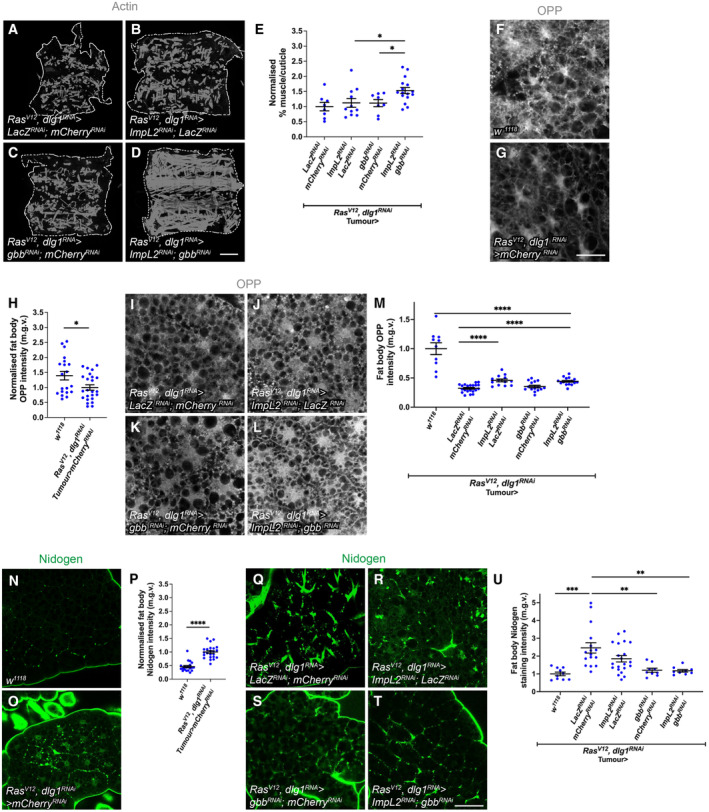
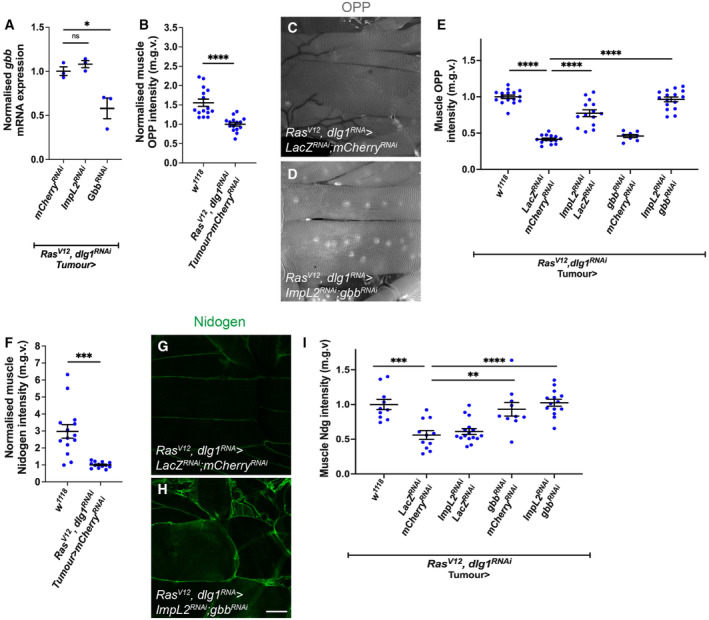
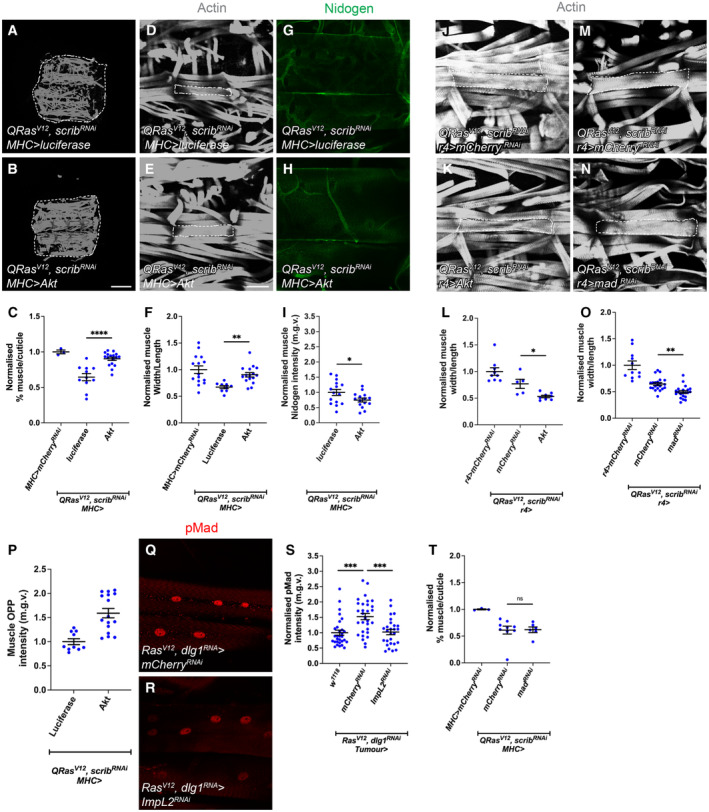
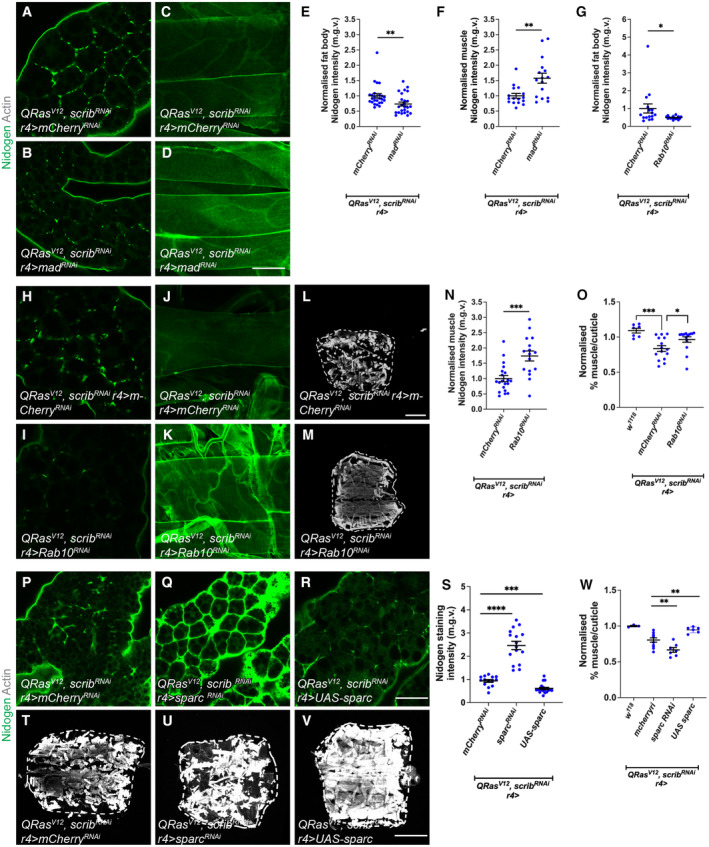
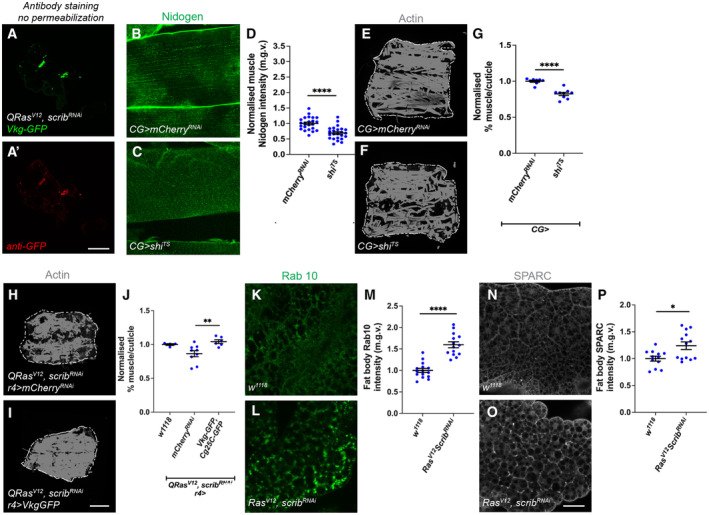
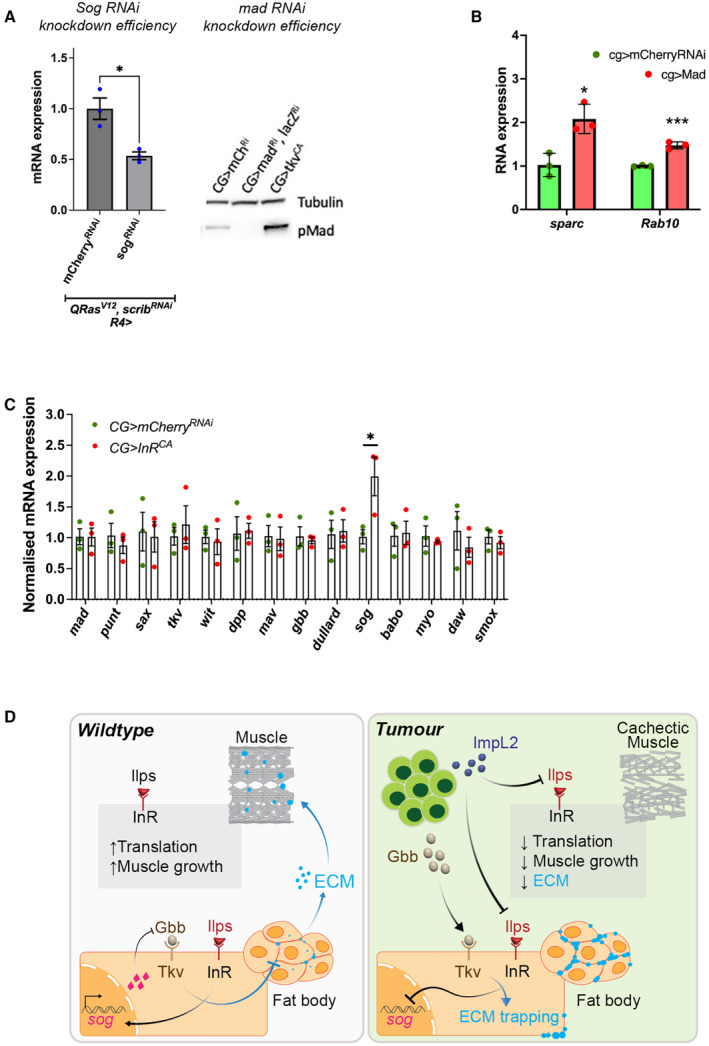
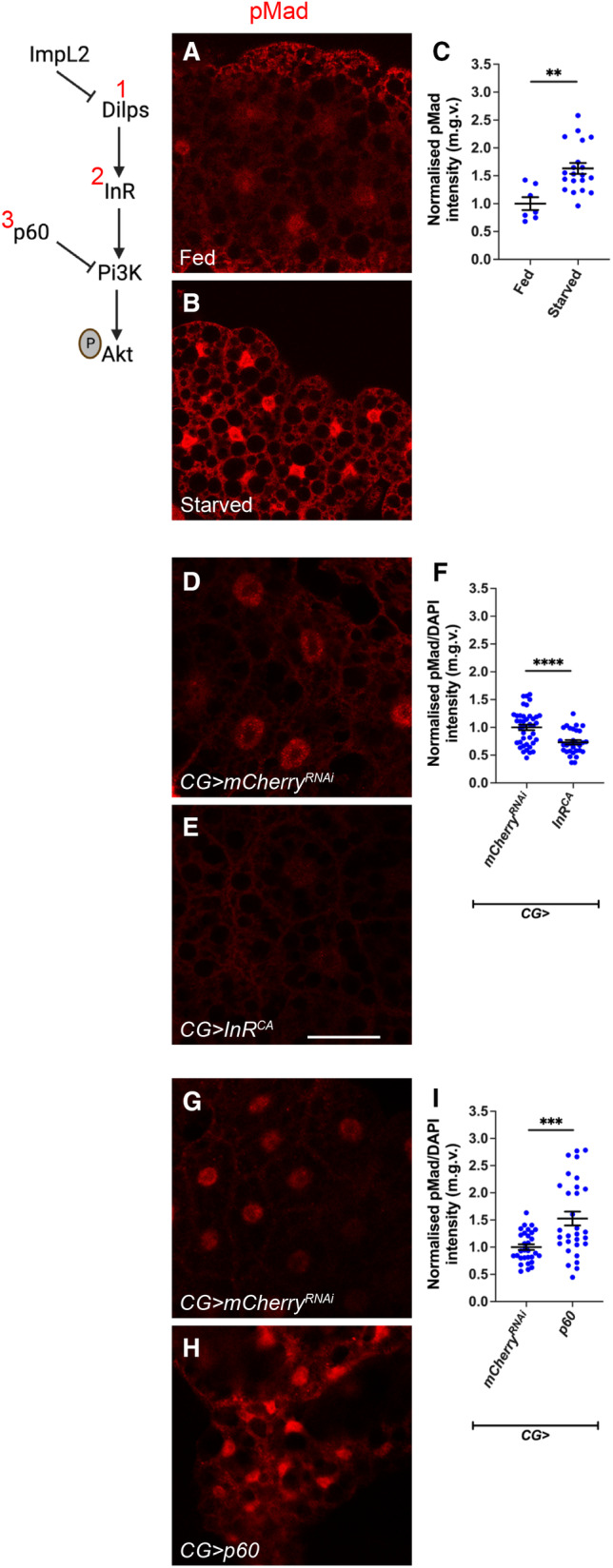
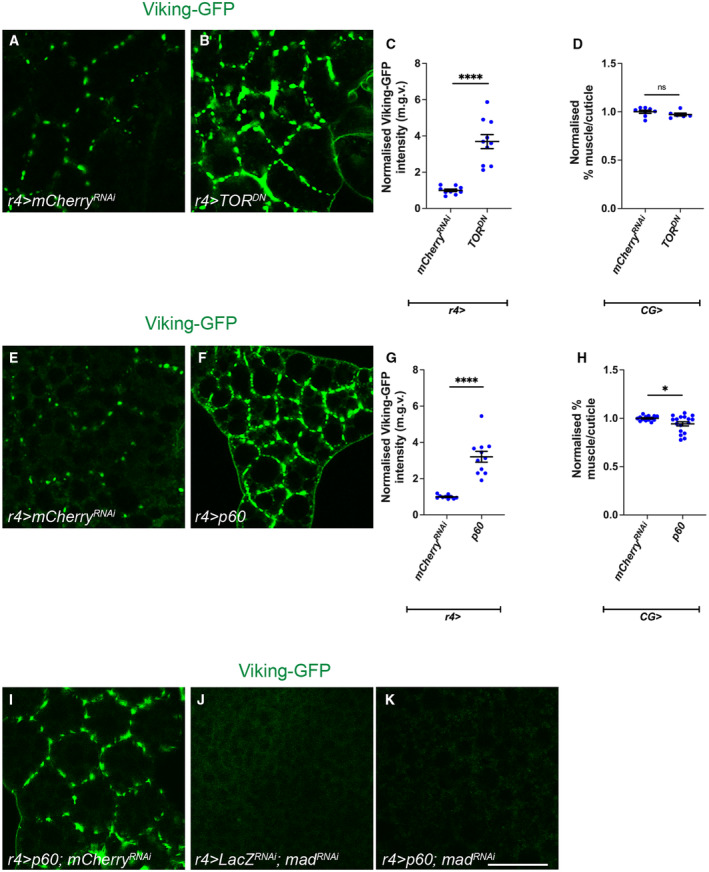
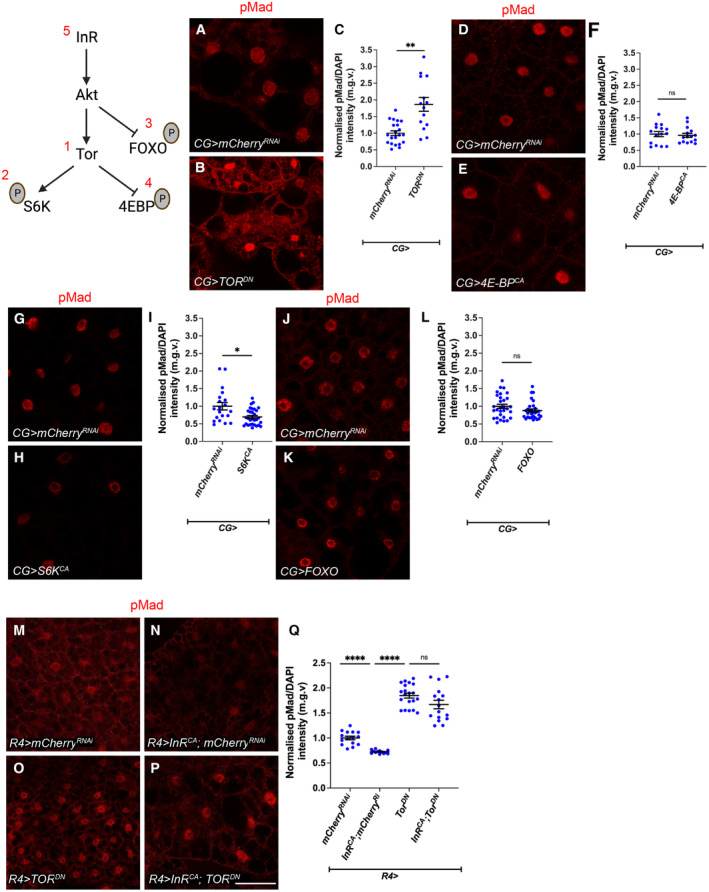
In this study, we found that in the adipose tissue of wildtype animals, insulin and TGF-β signalling converge via a BMP antagonist short gastrulation (sog) to regulate ECM remodelling. In tumour bearing animals, Sog also modulates TGF-β signalling to regulate ECM accumulation in the fat body. TGF-β signalling causes ECM retention in the fat body and subsequently depletes muscles of fat body-derived ECM proteins. Activation of insulin signalling, inhibition of TGF-β signalling, or modulation of ECM levels via SPARC, Rab10 or Collagen IV in the fat body, is able to rescue tissue wasting in the presence of tumour. Together, our study highlights the importance of adipose ECM remodelling in the context of cancer cachexia.
在这项研究中,我们发现,在野生型动物的脂肪组织中,胰岛素和 TGF-β 信号通过 BMP 拮抗剂短胚层(sog)汇聚,以调节细胞外基质重塑。在肿瘤荷瘤动物中,Sog 还调节 TGF-β 信号以调节脂肪体中的细胞外基质积累。TGF-β 信号导致脂肪体中细胞外基质的保留,随后耗尽肌肉中脂肪体衍生的细胞外基质蛋白。在存在肿瘤的情况下,通过脂肪体中的 SPARC、Rab10 或 Collagen IV 激活胰岛素信号、抑制 TGF-β 信号或调节细胞外基质水平,能够挽救组织消耗。总之,我们的研究强调了癌症恶病质背景下脂肪细胞外基质重塑的重要性。