Department of Physiology, Development and Neuroscience, University of Cambridge, Downing Site, Cambridge, CB2 3DY, UK.
Cambridge Advanced Imaging Centre, University of Cambridge, Downing Site, Cambridge, CB2 3DY, UK.
Epigenetics Chromatin. 2024 Mar 25;17(1):8. doi: 10.1186/s13072-024-00535-9.
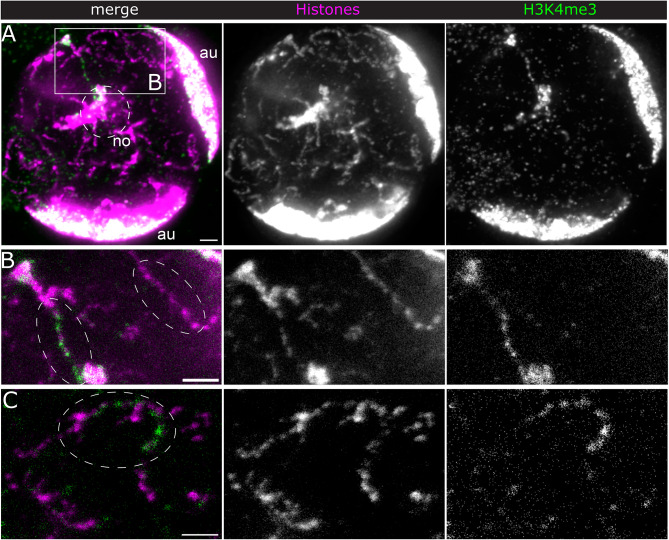
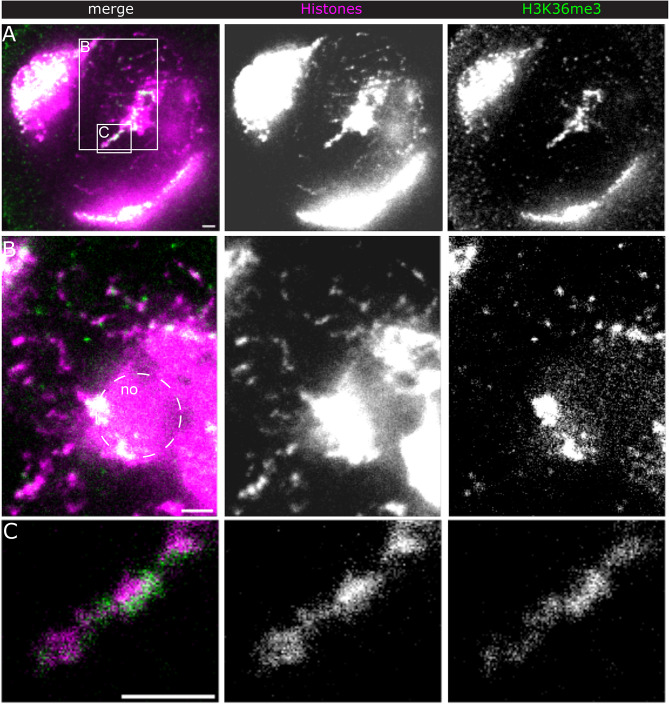
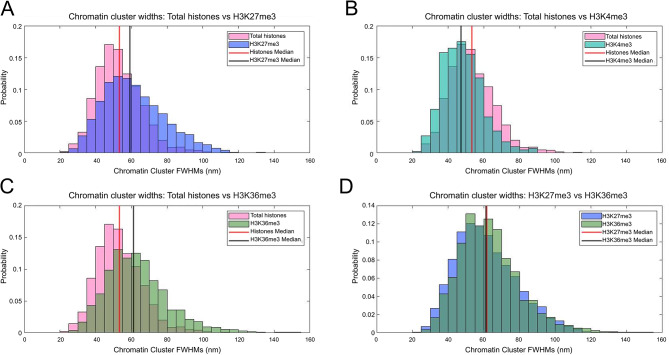
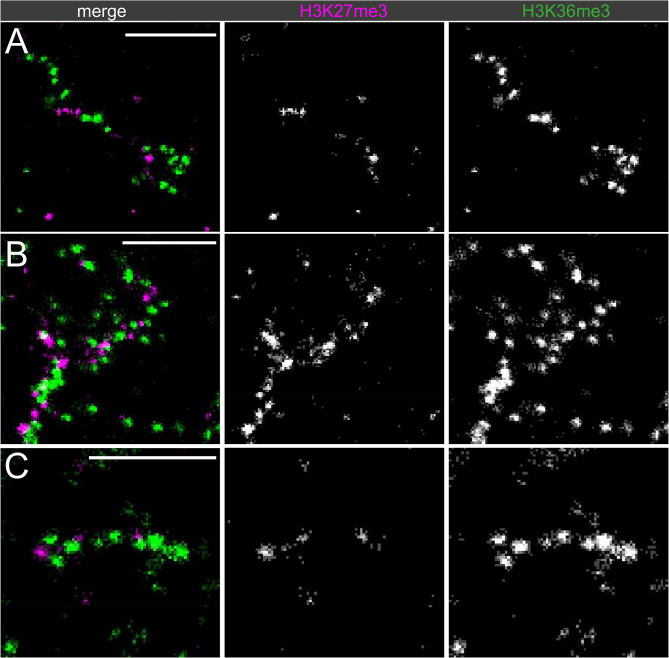
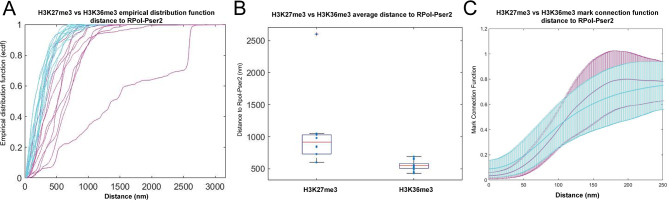
Chromatin state is thought to impart regulatory function to the underlying DNA sequence. This can be established through histone modifications and chromatin organisation, but exactly how these factors relate to one another to regulate gene expression is unclear. In this study, we have used super-resolution microscopy to image the Y loops of Drosophila melanogaster primary spermatocytes, which are enormous transcriptionally active chromatin fibres, each representing single transcription units that are individually resolvable in the nuclear interior. We previously found that the Y loops consist of regular clusters of nucleosomes, with an estimated median of 54 nucleosomes per cluster with wide variation.In this study, we report that the histone modifications H3K4me3, H3K27me3, and H3K36me3 are also clustered along the Y loops, with H3K4me3 more associated with diffuse chromatin compared to H3K27me3. These histone modifications form domains that can be stretches of Y loop chromatin micrometres long, or can be in short alternating domains. The different histone modifications are associated with different sizes of chromatin clusters and unique morphologies. Strikingly, a single chromatin cluster almost always only contains only one type of the histone modifications that were labelled, suggesting exclusivity, and therefore regulation at the level of individual chromatin clusters. The active mark H3K36me3 is more associated with actively elongating RNA polymerase II than H3K27me3, with polymerase often appearing on what are assumed to be looping regions on the periphery of chromatin clusters.These results provide a foundation for understanding the relationship between chromatin state, chromatin organisation, and transcription regulation - with potential implications for pause-release dynamics, splicing complex organisation and chromatin dynamics during polymerase progression along a gene.
染色质状态被认为赋予了底层 DNA 序列的调控功能。这可以通过组蛋白修饰和染色质组织来实现,但这些因素如何相互关联以调节基因表达还不清楚。在这项研究中,我们使用超分辨率显微镜来成像果蝇初级精母细胞的 Y 环,这些 Y 环是巨大的转录活性染色质纤维,每个纤维代表单个转录单位,在核内部可以单独分辨。我们之前发现 Y 环由规则的核小体簇组成,每个簇估计有 54 个核小体,变异范围很宽。在这项研究中,我们报告说组蛋白修饰 H3K4me3、H3K27me3 和 H3K36me3 也沿着 Y 环聚集,与 H3K27me3 相比,H3K4me3 更多地与弥散染色质相关。这些组蛋白修饰形成可以延伸数微米长的 Y 环染色质的域,或者可以形成短的交替域。不同的组蛋白修饰与不同大小的染色质簇和独特的形态相关。引人注目的是,单个染色质簇几乎总是只包含一种被标记的组蛋白修饰,这表明存在排他性,因此在单个染色质簇水平上进行调控。活性标记 H3K36me3 比 H3K27me3 更与正在延伸的 RNA 聚合酶 II 相关,聚合酶通常出现在染色质簇外围的假设的环化区域。这些结果为理解染色质状态、染色质组织和转录调控之间的关系提供了基础——这可能对暂停释放动力学、剪接复合物组织以及聚合酶沿着基因前进时的染色质动力学具有潜在影响。