Jaczynska Klaudia, Esser Victoria, Xu Junjie, Sari Levent, Lin Milo M, Rosenmund Christian, Rizo Josep
Department of Biophysics, University of Texas Southwestern Medical Center, Dallas, Texas 75390, USA.
Department of Biochemistry, University of Texas Southwestern Medical Center, Dallas, Texas 75390, USA.
bioRxiv. 2024 Jun 18:2024.06.17.599417. doi: 10.1101/2024.06.17.599417.
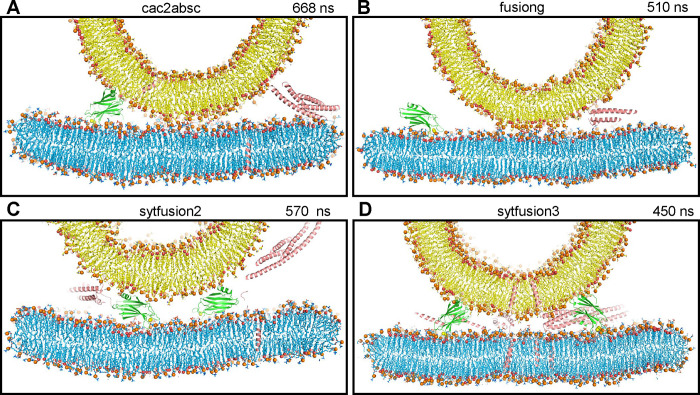
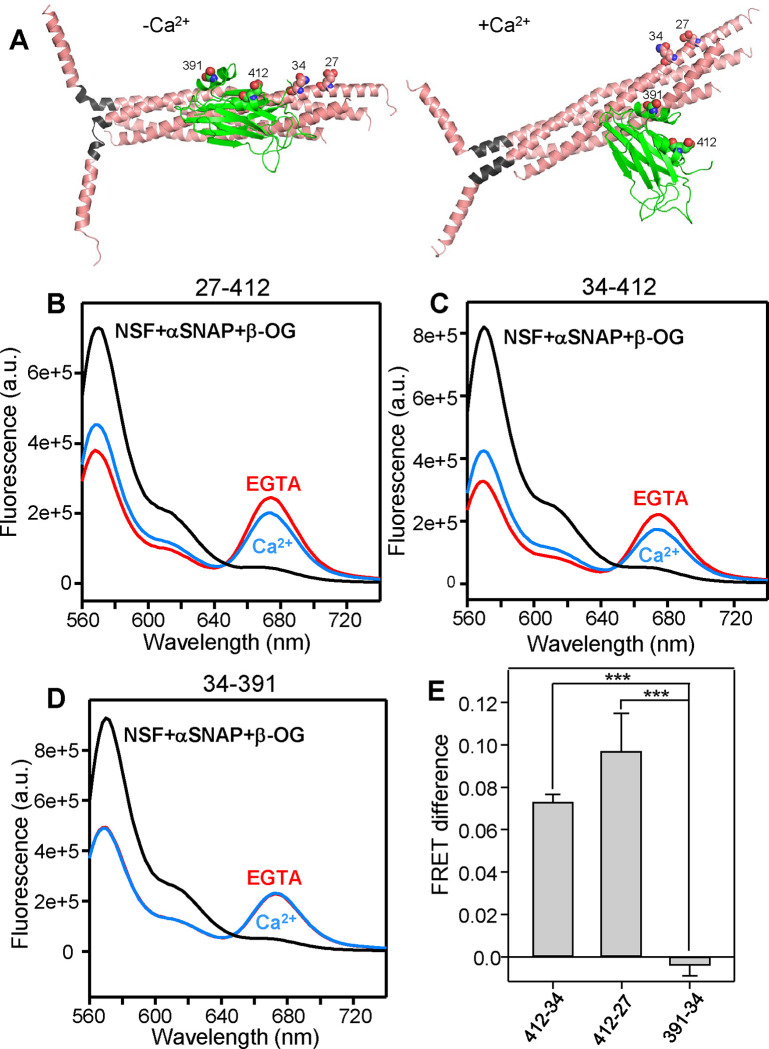
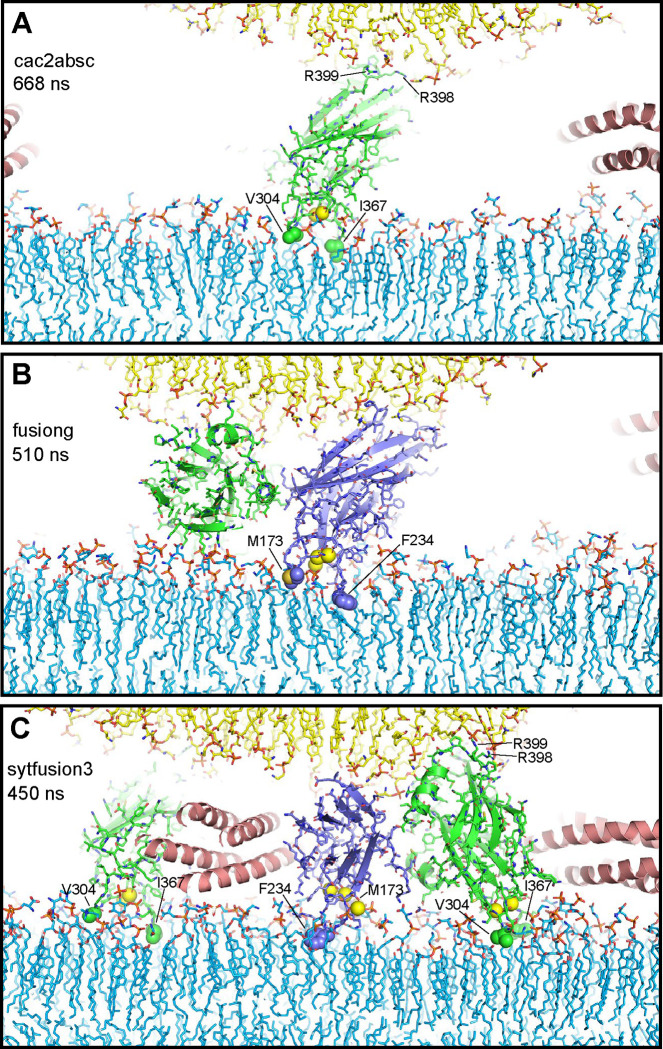
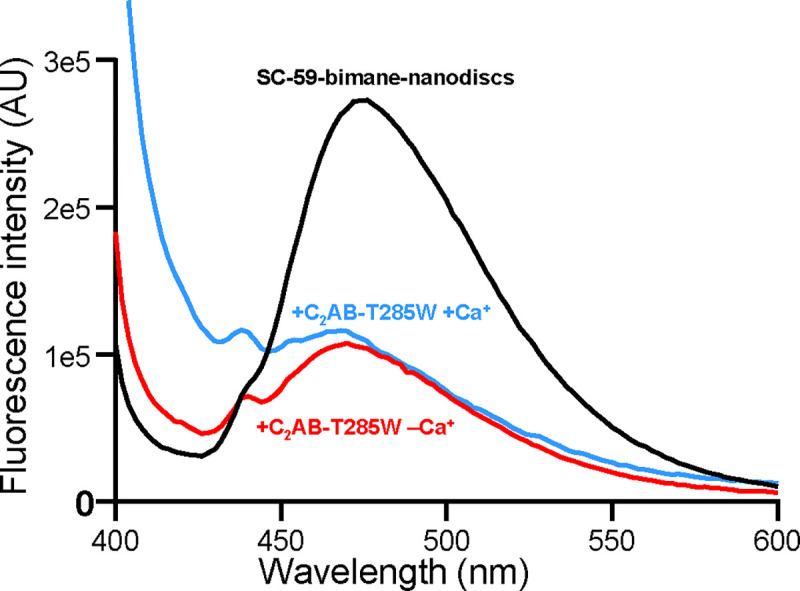
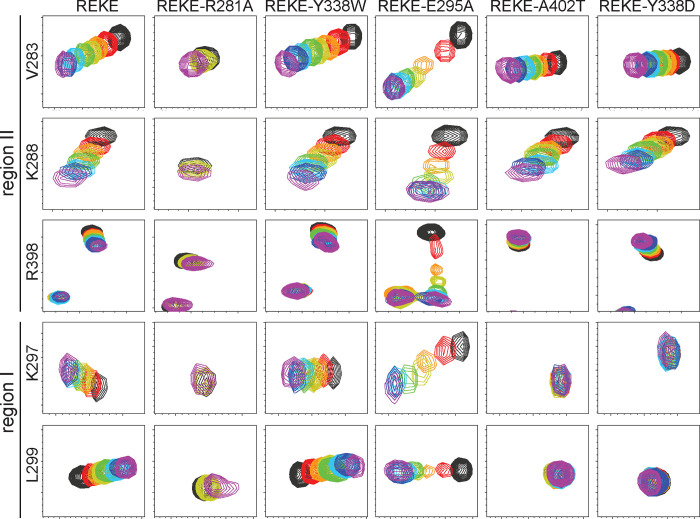
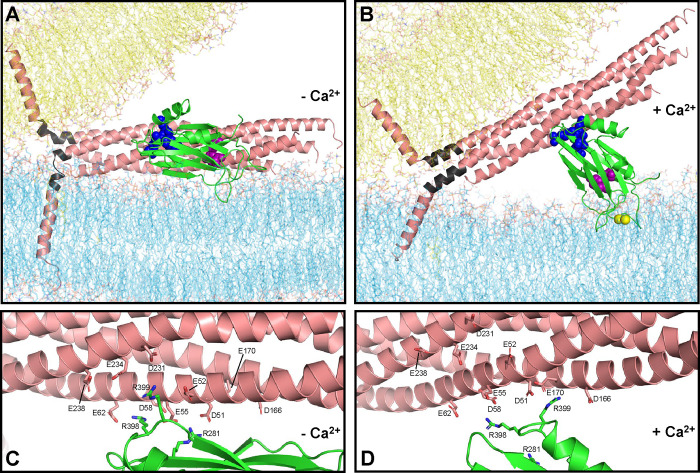
Neurotransmitter release is triggered in microseconds by Ca-binding to the Synaptotagmin-1 C domains and by SNARE complexes that form four-helix bundles between synaptic vesicles and plasma membranes, but the coupling mechanism between Ca-sensing and membrane fusion is unknown. Release requires extension of SNARE helices into juxtamembrane linkers that precede transmembrane regions (linker zippering) and binding of the Synaptotagmin-1 CB domain to SNARE complexes through a 'primary interface' comprising two regions (I and II). The Synaptotagmin-1 Ca-binding loops were believed to accelerate membrane fusion by inducing membrane curvature, perturbing lipid bilayers or helping bridge the membranes, but SNARE complex binding orients the Ca-binding loops away from the fusion site, hindering these putative activities. Molecular dynamics simulations now suggest that Synaptotagmin-1 C domains near the site of fusion hinder SNARE action, providing an explanation for this paradox and arguing against previous models of Sytnaptotagmin-1 action. NMR experiments reveal that binding of CB domain arginines to SNARE acidic residues at region II remains after disruption of region I. These results and fluorescence resonance energy transfer assays, together with previous data, suggest that Ca causes reorientation of the CB domain on the membrane and dissociation from the SNAREs at region I but not region II. Based on these results and molecular modeling, we propose that Synaptotagmin-1 acts as a lever that pulls the SNARE complex when Ca causes reorientation of the CB domain, facilitating linker zippering and fast membrane fusion. This hypothesis is supported by the electrophysiological data described in the accompanying paper.
钙离子与突触结合蛋白-1的C结构域结合以及SNARE复合体在突触小泡和质膜之间形成四螺旋束,在微秒内触发神经递质释放,但钙离子感应与膜融合之间的偶联机制尚不清楚。释放需要SNARE螺旋延伸到跨膜区域之前的近膜连接子中(连接子拉链化),以及突触结合蛋白-1的CB结构域通过包含两个区域(I和II)的“主要界面”与SNARE复合体结合。人们认为突触结合蛋白-1的钙离子结合环通过诱导膜曲率、扰动脂质双层或帮助连接膜来加速膜融合,但SNARE复合体结合使钙离子结合环远离融合位点,阻碍了这些假定的活动。分子动力学模拟现在表明,融合位点附近的突触结合蛋白-1的C结构域阻碍SNARE作用,为这一矛盾提供了解释,并与之前关于突触结合蛋白-1作用的模型相悖。核磁共振实验表明,在区域I被破坏后,CB结构域的精氨酸与区域II的SNARE酸性残基的结合仍然存在。这些结果和荧光共振能量转移分析,以及之前的数据表明,钙离子导致CB结构域在膜上重新定向,并在区域I而非区域II与SNARE解离。基于这些结果和分子建模,我们提出,当钙离子导致CB结构域重新定向时,突触结合蛋白-1作为一个杠杆拉动SNARE复合体,促进连接子拉链化和快速膜融合。这一假设得到了随附论文中描述的电生理数据的支持。