Ono Kazuya, Jarysta Amandine, Hughes Natasha C, Jukic Alma, Chang Hui Ho Vanessa, Deans Michael R, Eatock Ruth Anne, Cullen Kathleen E, Kindt Katie, Tarchini Basile
Department of Neurobiology, University of Chicago, Chicago, IL, 60637, USA.
The Jackson Laboratory, Bar Harbor, ME 04609, USA.
bioRxiv. 2024 Sep 6:2024.03.26.586740. doi: 10.1101/2024.03.26.586740.
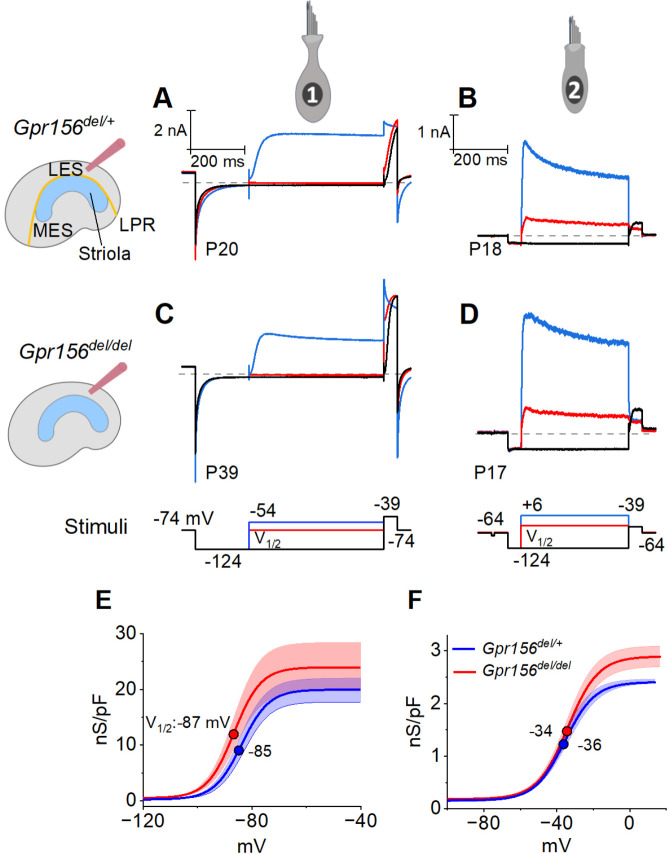
Otolith organs in the inner ear and neuromasts in the fish lateral-line harbor two populations of hair cells oriented to detect stimuli in opposing directions. The underlying mechanism is highly conserved: the transcription factor EMX2 is regionally expressed in just one hair cell population and acts through the receptor GPR156 to reverse cell orientation relative to the other population. In mouse and zebrafish, loss of Emx2 results in sensory organs that harbor only one hair cell orientation and are not innervated properly. In zebrafish, Emx2 also confers hair cells with reduced mechanosensory properties. Here, we leverage mouse and zebrafish models lacking GPR156 to determine how detecting stimuli of opposing directions serves vestibular function, and whether GPR156 has other roles besides orienting hair cells. We find that otolith organs in mouse mutants have normal zonal organization and normal type I-II hair cell distribution and mechano-electrical transduction properties. In contrast, zebrafish mutants lack the smaller mechanically-evoked signals that characterize Emx2-positive hair cells. Loss of GPR156 does not affect orientation-selectivity of afferents in mouse utricle or zebrafish neuromasts. Consistent with normal otolith organ anatomy and afferent selectivity, mutant mice do not show overt vestibular dysfunction. Instead, performance on two tests that engage otolith organs is significantly altered - swimming and off-vertical-axis rotation. We conclude that GPR156 relays hair cell orientation and transduction information downstream of EMX2, but not selectivity for direction-specific afferents. These results clarify how molecular mechanisms that confer bi-directionality to sensory organs contribute to function, from single hair cell physiology to animal behavior.
内耳中的耳石器官以及鱼类侧线中的神经丘含有两类毛细胞,它们的方向相反,用于检测不同方向的刺激。其潜在机制高度保守:转录因子EMX2仅在一类毛细胞中呈区域表达,并通过受体GPR156发挥作用,使这类毛细胞相对于另一类毛细胞的方向发生反转。在小鼠和斑马鱼中,Emx2缺失会导致感觉器官中仅存在一种毛细胞方向,且神经支配异常。在斑马鱼中,Emx2还会使毛细胞的机械感觉特性降低。在此,我们利用缺乏GPR156的小鼠和斑马鱼模型,来确定检测相反方向的刺激如何发挥前庭功能,以及GPR156除了使毛细胞定向之外是否还有其他作用。我们发现,小鼠突变体中的耳石器官具有正常的区域组织、正常的I型-II型毛细胞分布以及机械电转换特性。相比之下,斑马鱼突变体缺乏表征Emx2阳性毛细胞的较小的机械诱发信号。GPR156的缺失并不影响小鼠椭圆囊或斑马鱼神经丘中传入神经的方向选择性。与正常的耳石器官解剖结构和传入神经选择性一致,突变小鼠未表现出明显的前庭功能障碍。相反,在两项涉及耳石器官的测试——游泳和离垂直轴旋转中的表现却发生了显著改变。我们得出结论,GPR156在内耳毛细胞方向和转导信息传递过程中发挥作用,但是不参与方向特异性传入神经的方向选择性。这些结果阐明了赋予感觉器官双向性的分子机制是如何从单个毛细胞生理功能到动物行为,进而对整体功能产生影响的。