The Solomon H. Snyder Department of Neuroscience, Johns Hopkins University School of Medicine, Baltimore, MD, USA.
Kavli Neuroscience Discovery Institute, Johns Hopkins University, Baltimore, MD, USA.
Nature. 2024 Nov;635(8038):398-405. doi: 10.1038/s41586-024-08027-2. Epub 2024 Oct 2.
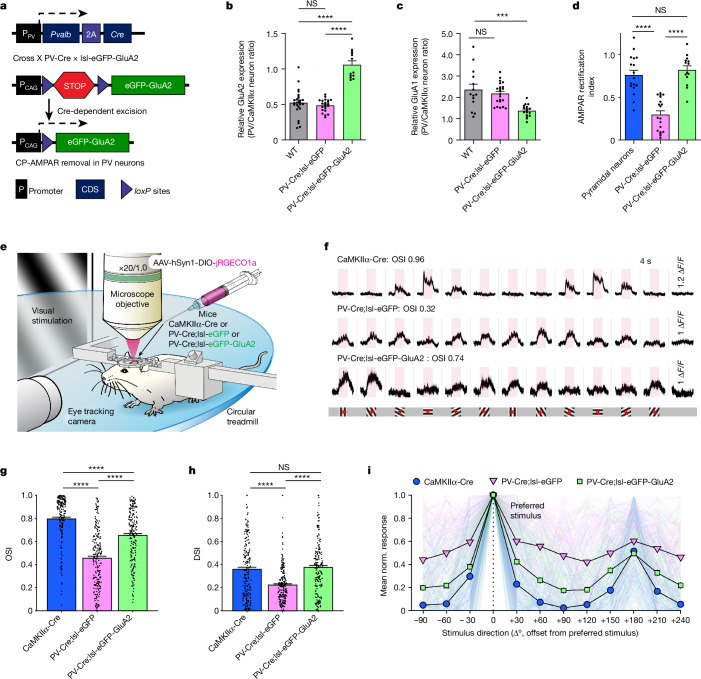
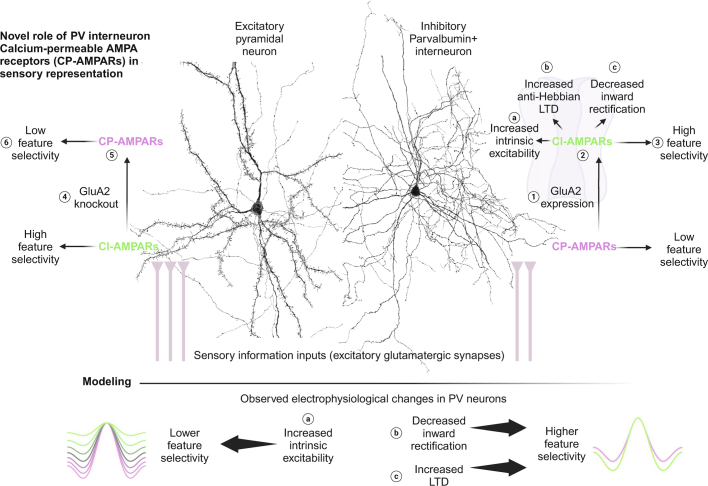
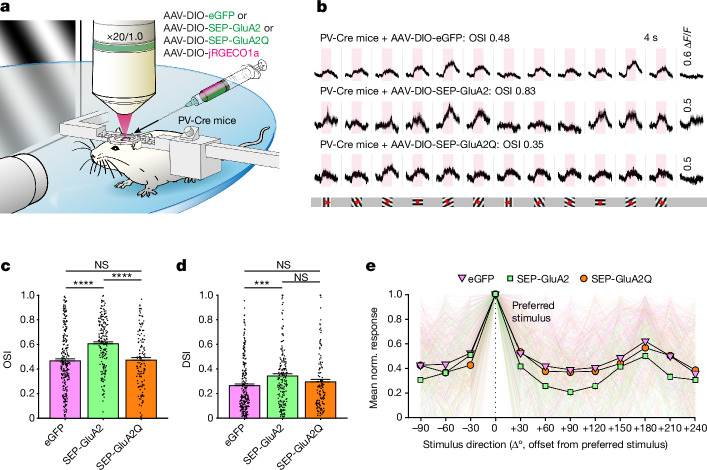
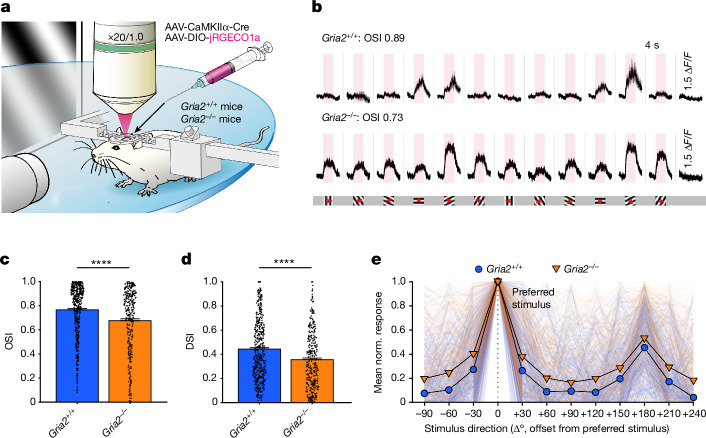
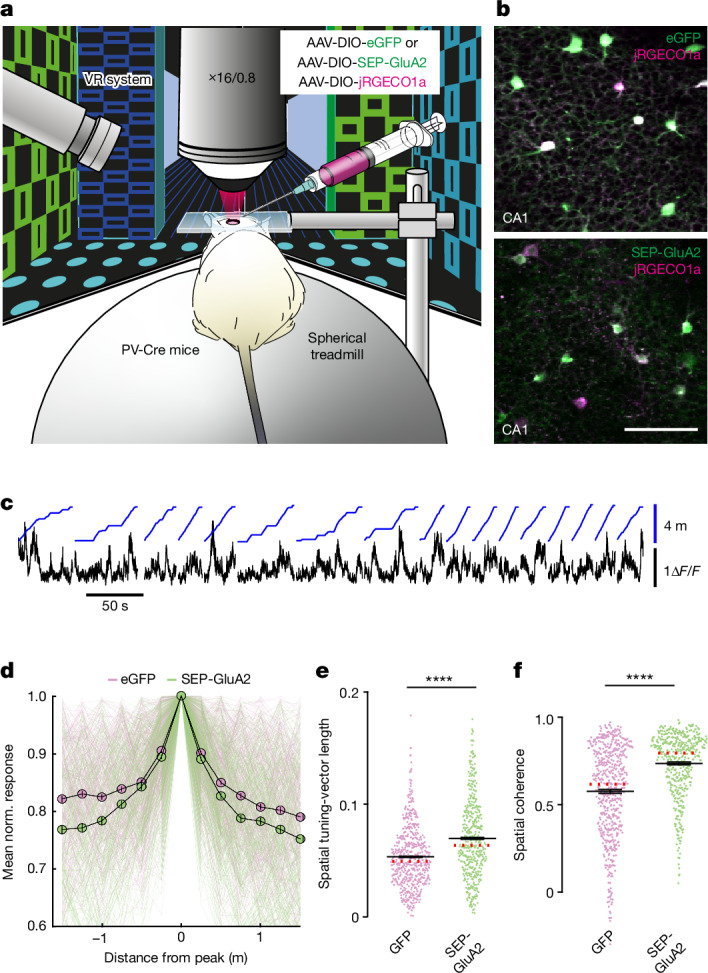
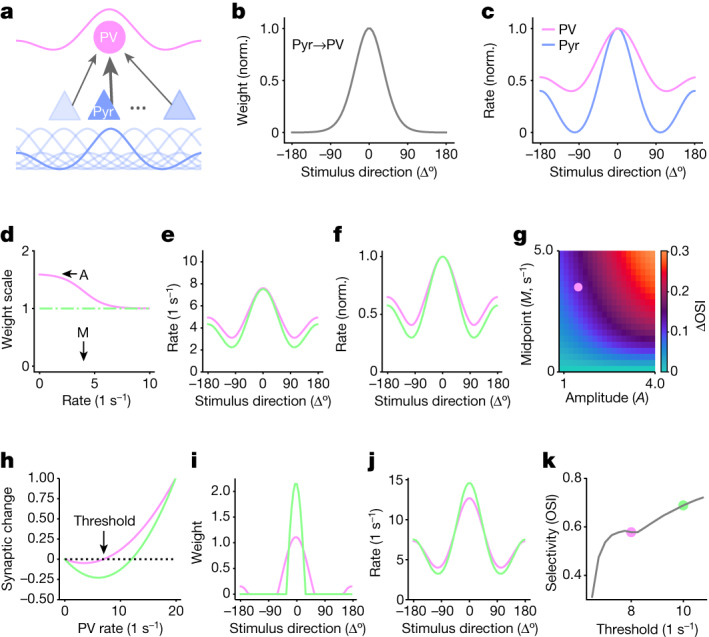
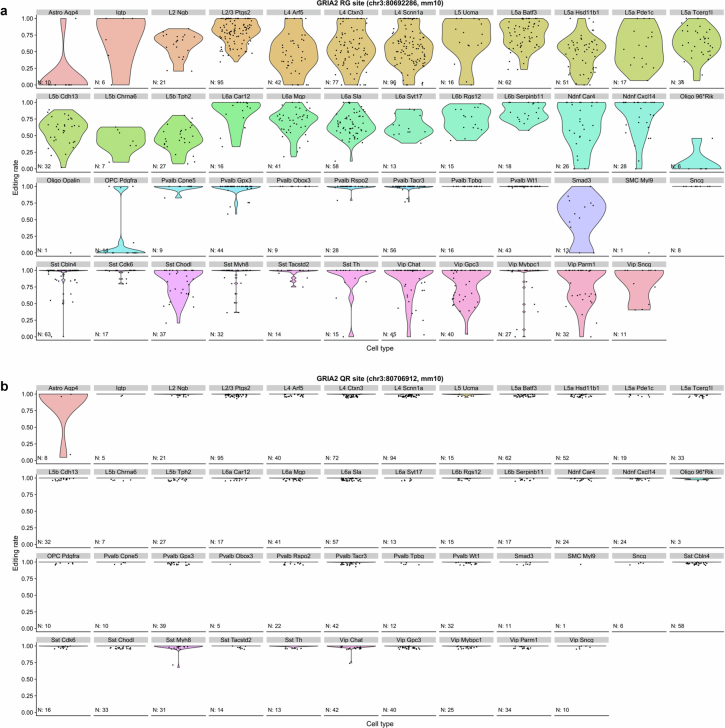
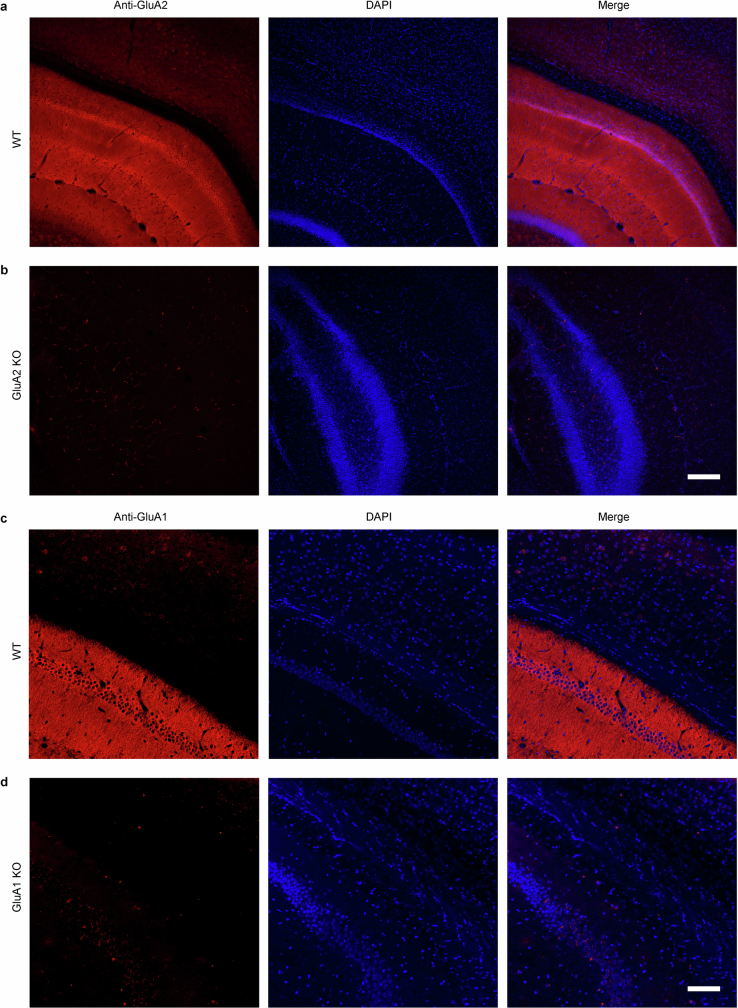
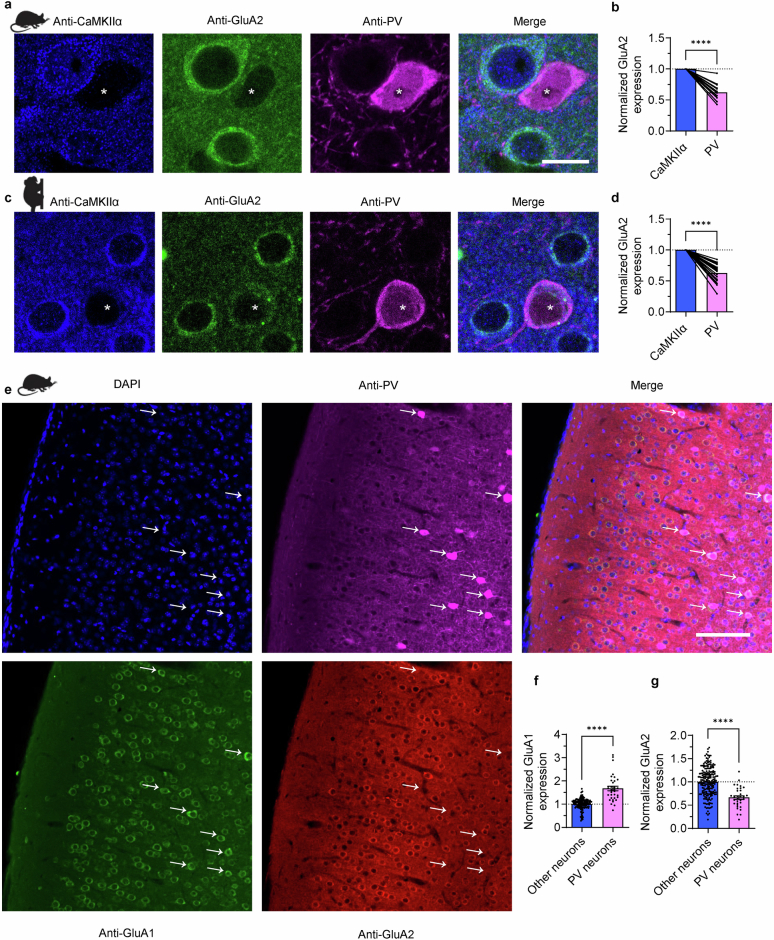
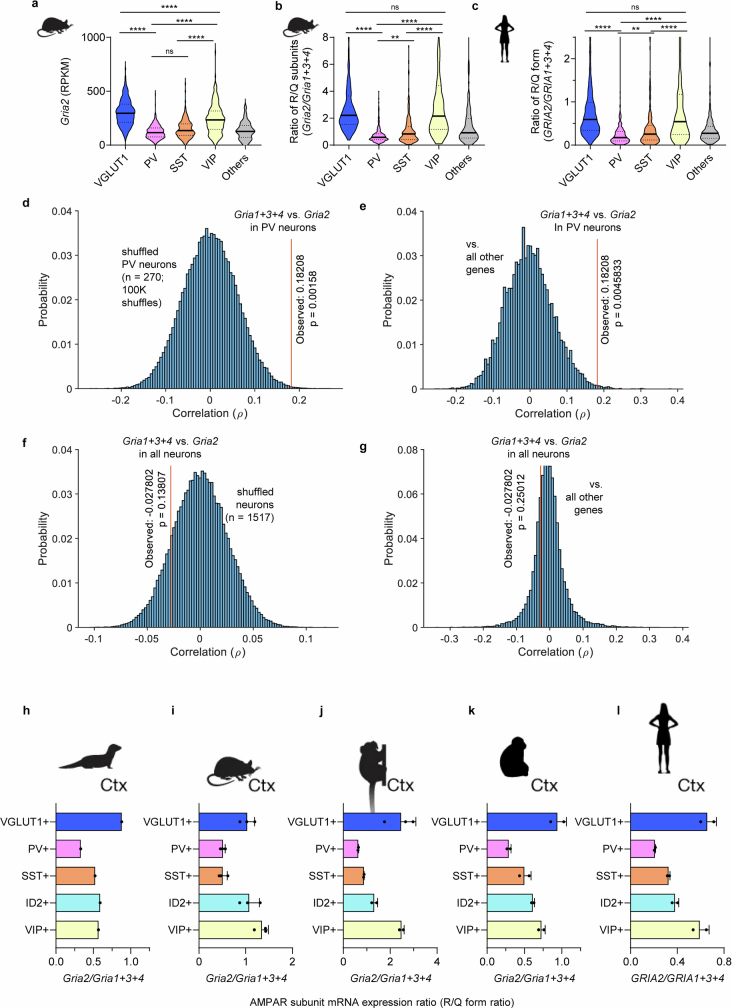
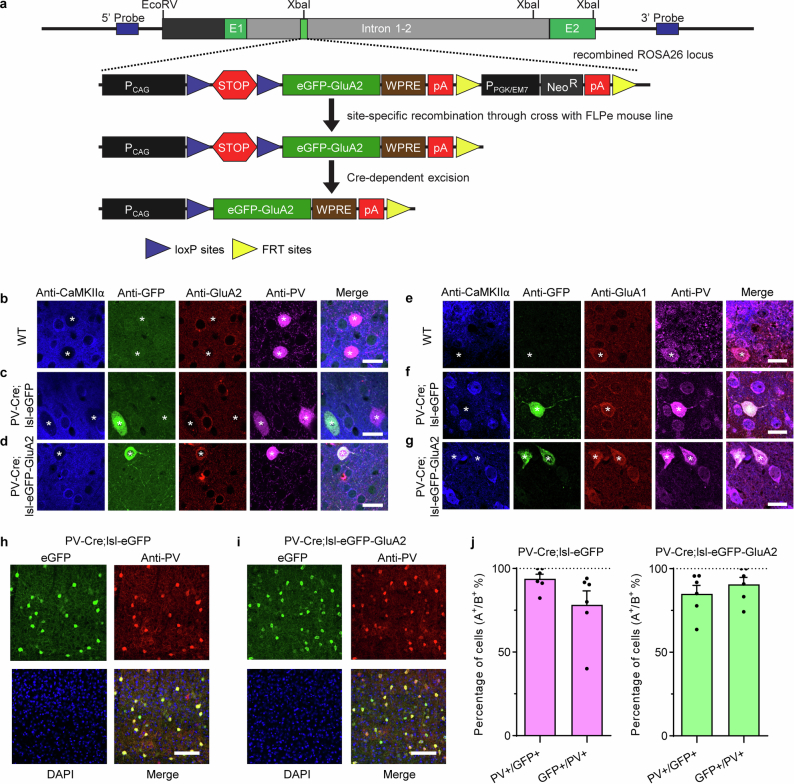
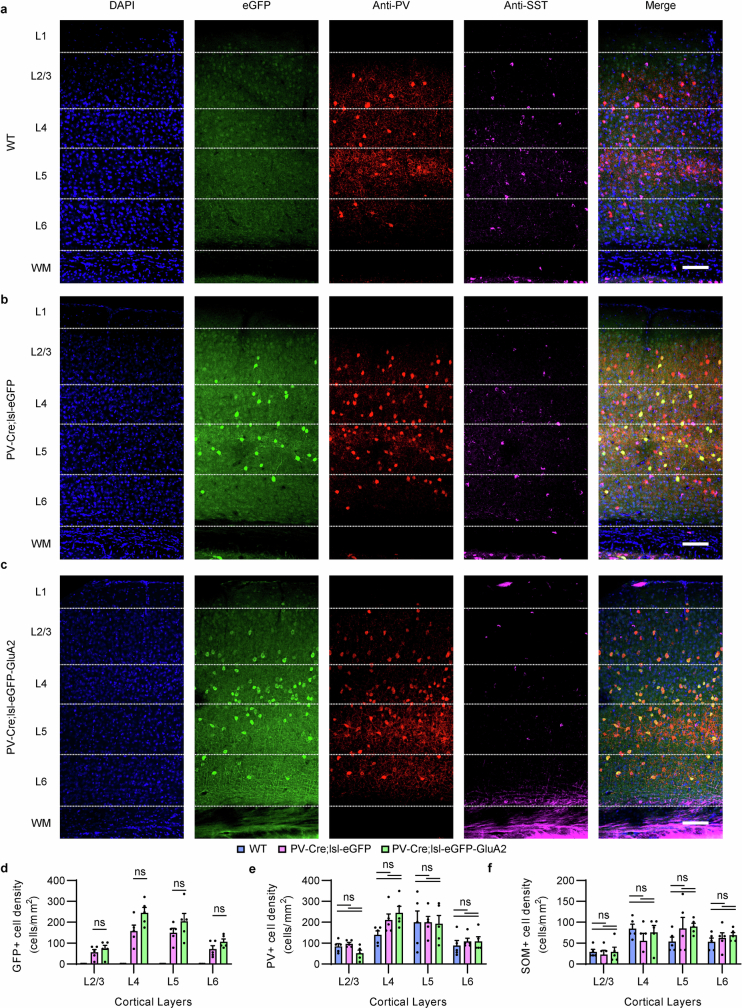
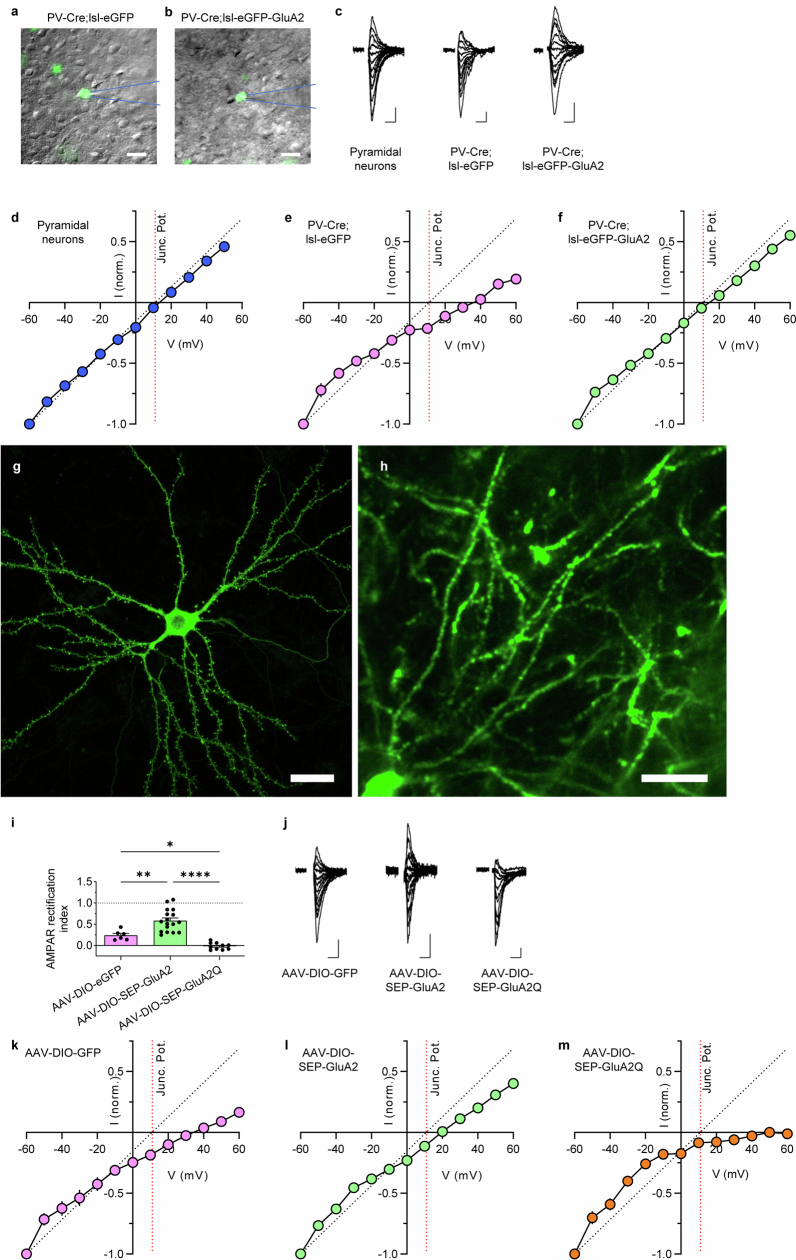
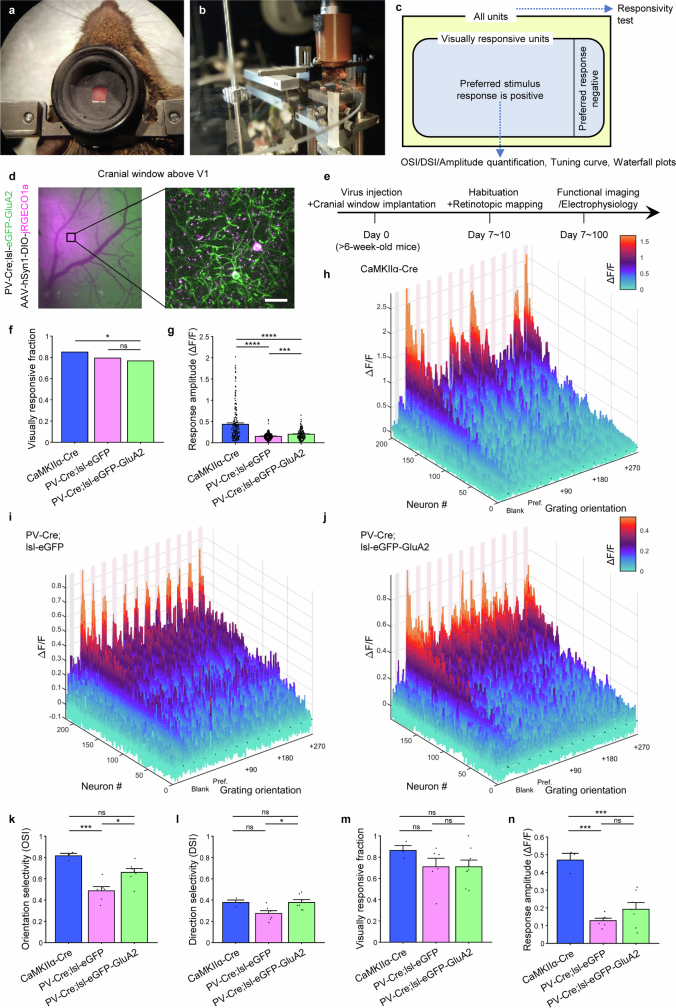
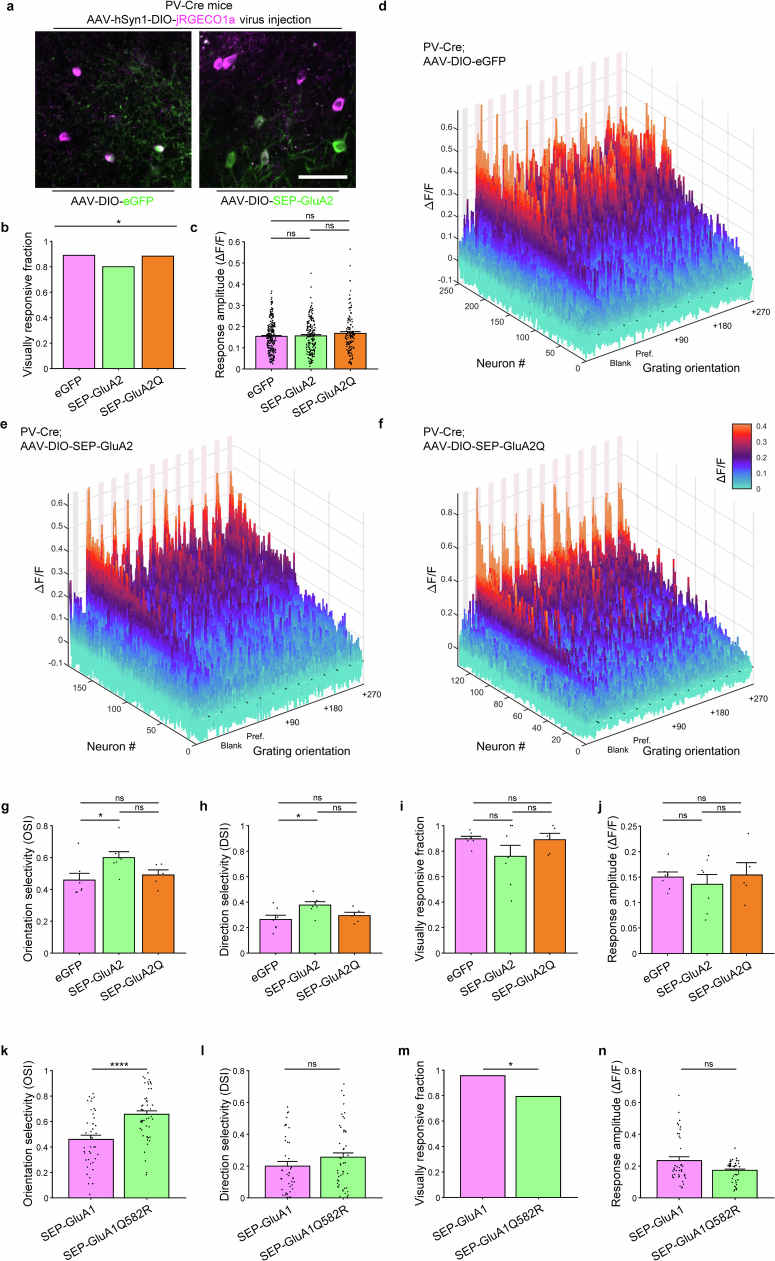
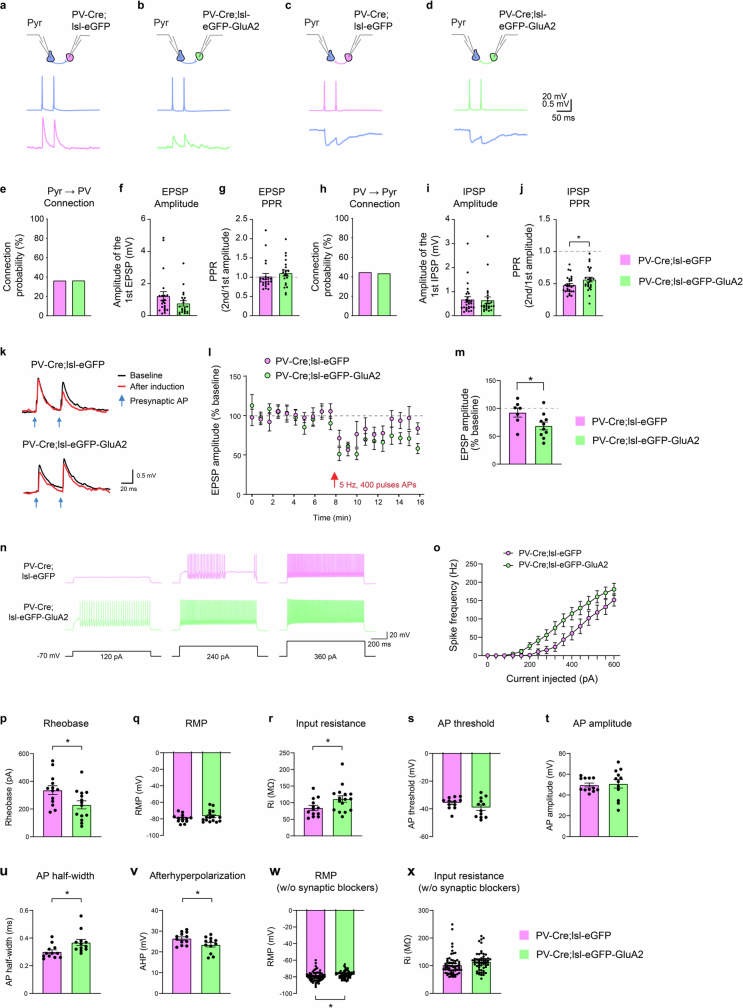
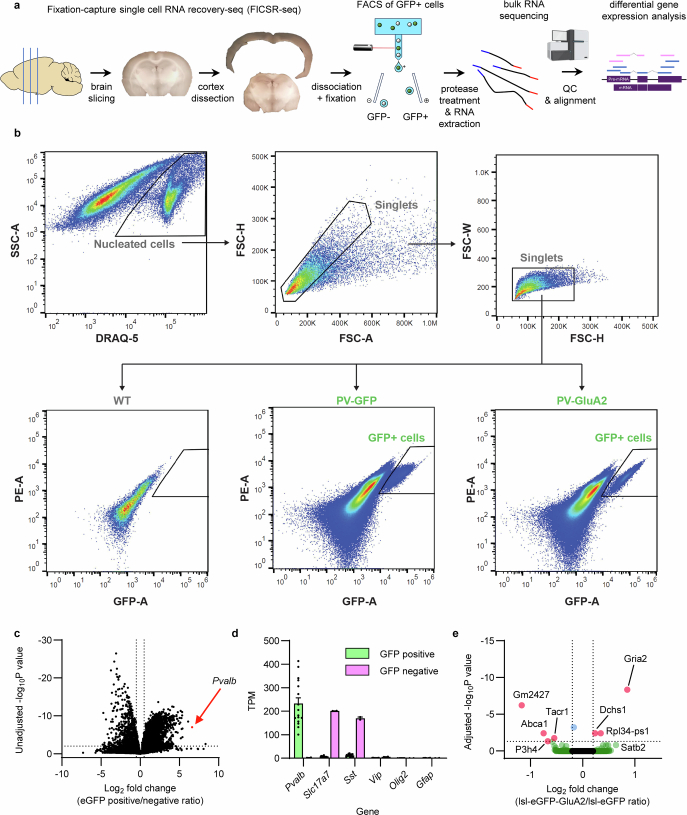
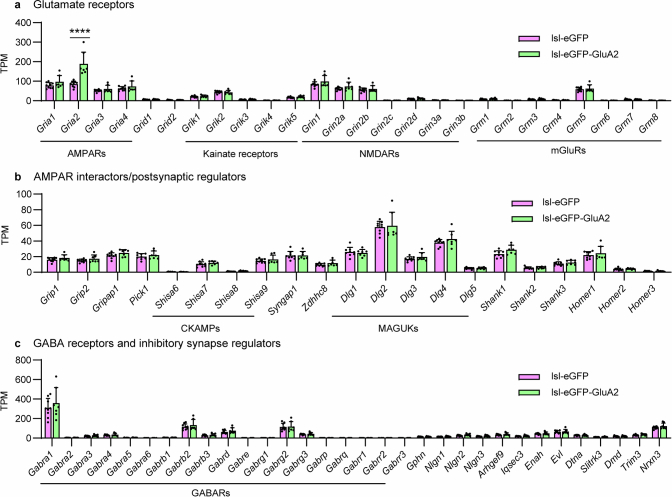
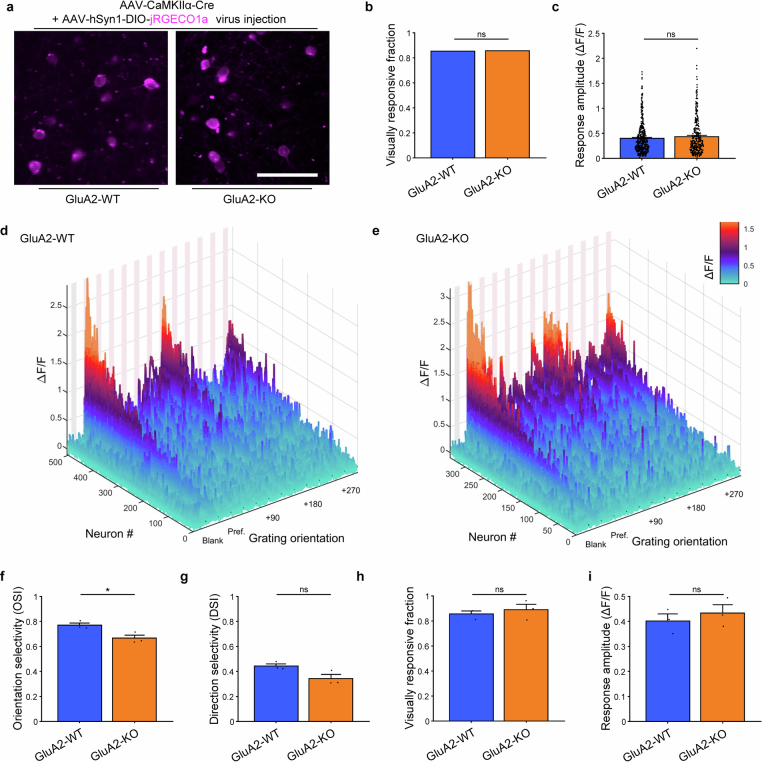
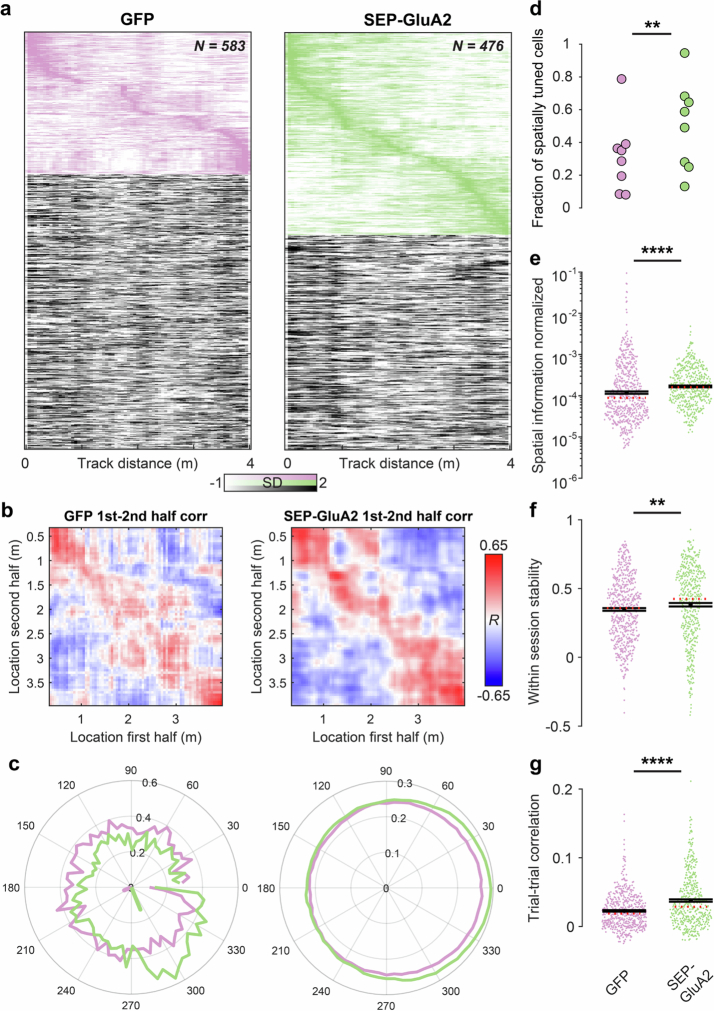
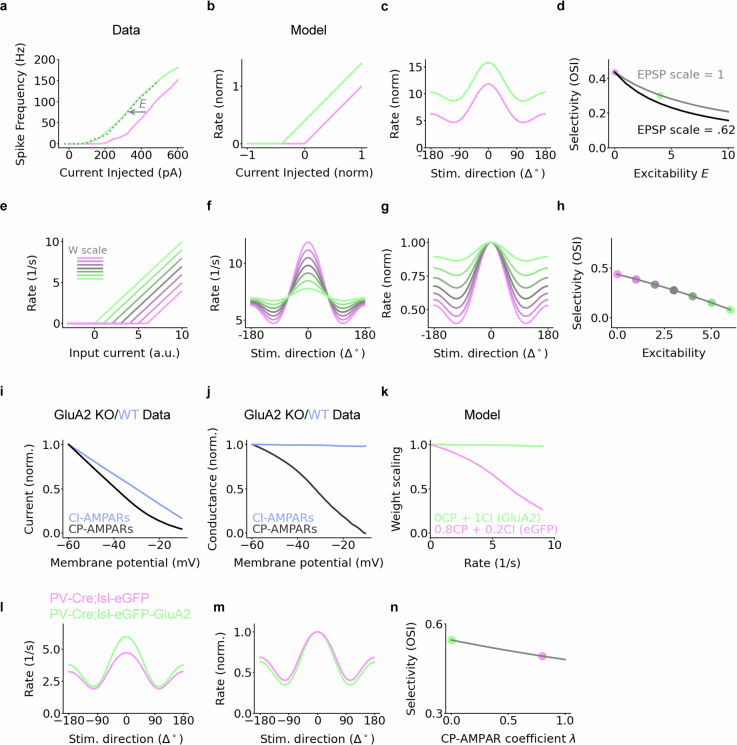
The brain helps us survive by forming internal representations of the external world. Excitatory cortical neurons are often precisely tuned to specific external stimuli. However, inhibitory neurons, such as parvalbumin-positive (PV) interneurons, are generally less selective. PV interneurons differ from excitatory neurons in their neurotransmitter receptor subtypes, including AMPA (α-amino-3-hydroxy-5-methyl-4-isoxazole propionic acid) receptors (AMPARs). Excitatory neurons express calcium-impermeable AMPARs that contain the GluA2 subunit (encoded by GRIA2), whereas PV interneurons express receptors that lack the GluA2 subunit and are calcium-permeable (CP-AMPARs). Here we demonstrate a causal relationship between CP-AMPAR expression and the low feature selectivity of PV interneurons. We find low expression stoichiometry of GRIA2 mRNA relative to other subunits in PV interneurons that is conserved across ferrets, rodents, marmosets and humans, and causes abundant CP-AMPAR expression. Replacing CP-AMPARs in PV interneurons with calcium-impermeable AMPARs increased their orientation selectivity in the visual cortex. Manipulations to induce sparse CP-AMPAR expression demonstrated that this increase was cell-autonomous and could occur with changes beyond development. Notably, excitatory-PV interneuron connectivity rates and unitary synaptic strength were unaltered by CP-AMPAR removal, which suggested that the selectivity of PV interneurons can be altered without markedly changing connectivity. In Gria2-knockout mice, in which all AMPARs are calcium-permeable, excitatory neurons showed significantly degraded orientation selectivity, which suggested that CP-AMPARs are sufficient to drive lower selectivity regardless of cell type. Moreover, hippocampal PV interneurons, which usually exhibit low spatial tuning, became more spatially selective after removing CP-AMPARs, which indicated that CP-AMPARs suppress the feature selectivity of PV interneurons independent of modality. These results reveal a new role of CP-AMPARs in maintaining low-selectivity sensory representation in PV interneurons and implicate a conserved molecular mechanism that distinguishes this cell type in the neocortex.
大脑通过形成对外界的内部表示来帮助我们生存。兴奋性皮质神经元通常对特定的外部刺激精确调谐。然而,抑制性神经元,如钙结合蛋白阳性(PV)中间神经元,通常选择性较低。PV 中间神经元在其神经递质受体亚型上与兴奋性神经元不同,包括 AMPA(α-氨基-3-羟基-5-甲基-4-异恶唑丙酸)受体(AMPAR)。兴奋性神经元表达钙不可渗透的 AMPAR,其包含 GluA2 亚基(由 GRIA2 编码),而 PV 中间神经元表达缺乏 GluA2 亚基且钙可渗透的受体(CP-AMPAR)。在这里,我们证明了 CP-AMPAR 表达与 PV 中间神经元的低特征选择性之间存在因果关系。我们发现,在 PV 中间神经元中,GRIA2 mRNA 的表达量相对于其他亚基相对较低,这种情况在雪貂、啮齿动物、狨猴和人类中是保守的,并且导致大量 CP-AMPAR 的表达。用钙不可渗透的 AMPAR 替换 PV 中间神经元中的 CP-AMPAR,增加了它们在视觉皮层中的方向选择性。诱导稀疏 CP-AMPAR 表达的操作表明,这种增加是细胞自主性的,并且可以在发育之外发生变化。值得注意的是,CP-AMPAR 去除后,兴奋性-PV 中间神经元连接率和单位突触强度没有改变,这表明 PV 中间神经元的选择性可以改变而不会显著改变连接。在所有 AMPAR 均为钙可渗透的 Gria2 敲除小鼠中,兴奋性神经元的方向选择性显著降低,这表明 CP-AMPAR 足以驱动较低的选择性,而与细胞类型无关。此外,通常表现出低空间调谐的海马 PV 中间神经元在去除 CP-AMPAR 后变得更具空间选择性,这表明 CP-AMPAR 独立于模态抑制 PV 中间神经元的特征选择性。这些结果揭示了 CP-AMPAR 在维持 PV 中间神经元低选择性感觉表示中的新作用,并暗示了一种保守的分子机制,该机制区分了新皮层中的这种细胞类型。