Liu Min, Yue Yanzhu, Chen Xubin, Xian Kexin, Dong Chao, Shi Ming, Xiong Haiqing, Tian Kang, Li Yuzhe, Zhang Qiangfeng Cliff, He Aibin
Institute of Molecular Medicine and National Biomedical Imaging Center, College of Future Technology, Peking-Tsinghua Center for Life Sciences and State Key Laboratory of Gene Function and Modulation Research, Peking University, Beijing, China.
Department of Cell Fate and Diseases, Jilin Provincial Key Laboratory of Women's Reproductive Health, Jilin Provincial Clinical Research Center for Birth Defect and Rare Disease, The First Hospital of Jilin University, Changchun, China.
Nature. 2025 Apr;640(8059):828-839. doi: 10.1038/s41586-025-08656-1. Epub 2025 Feb 26.
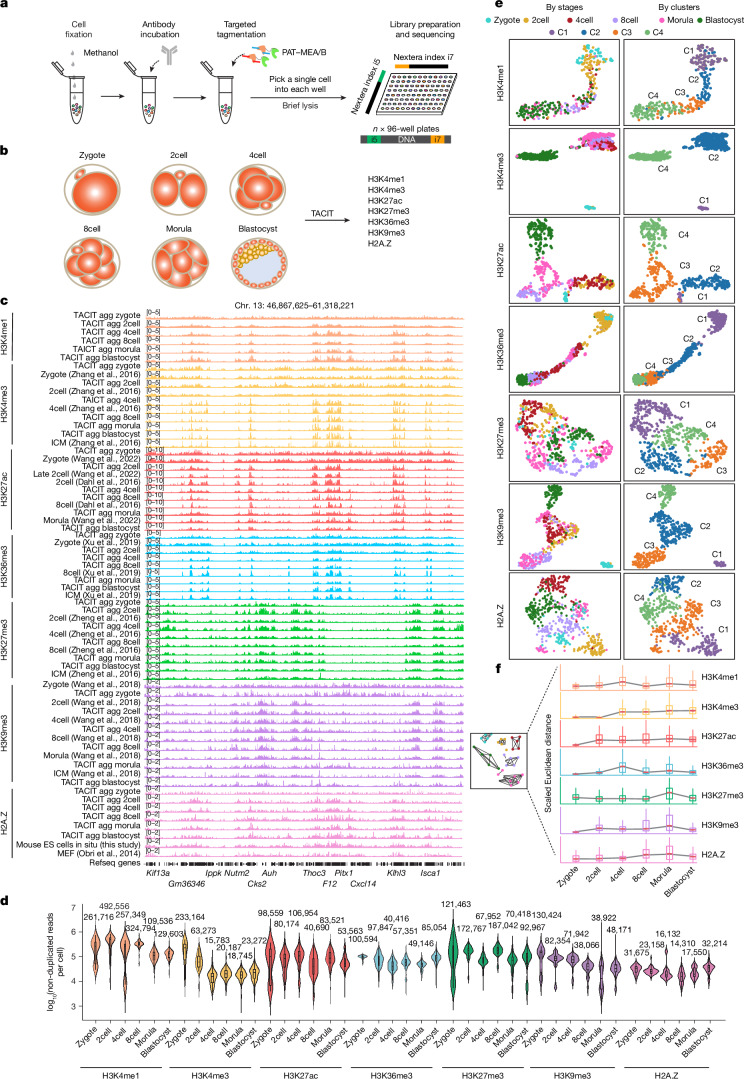
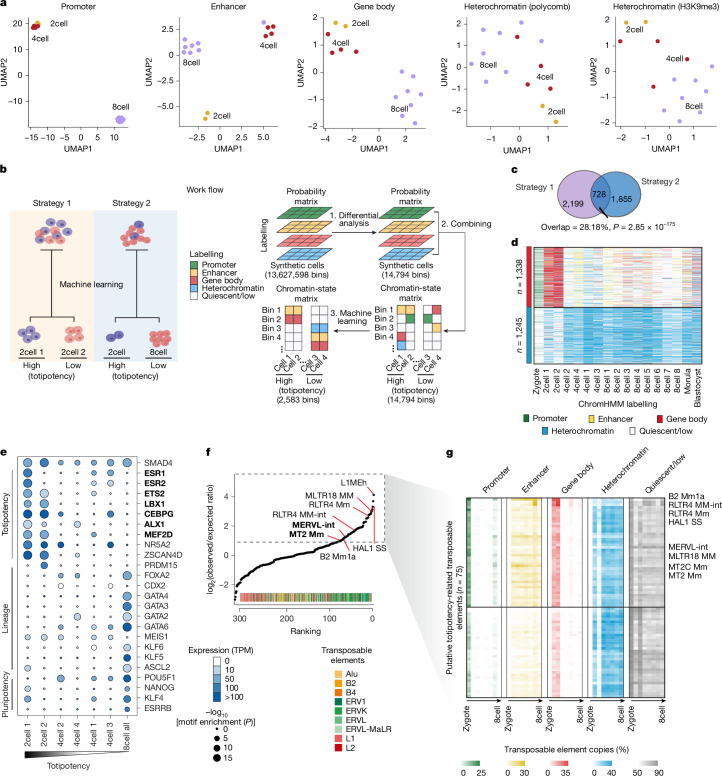
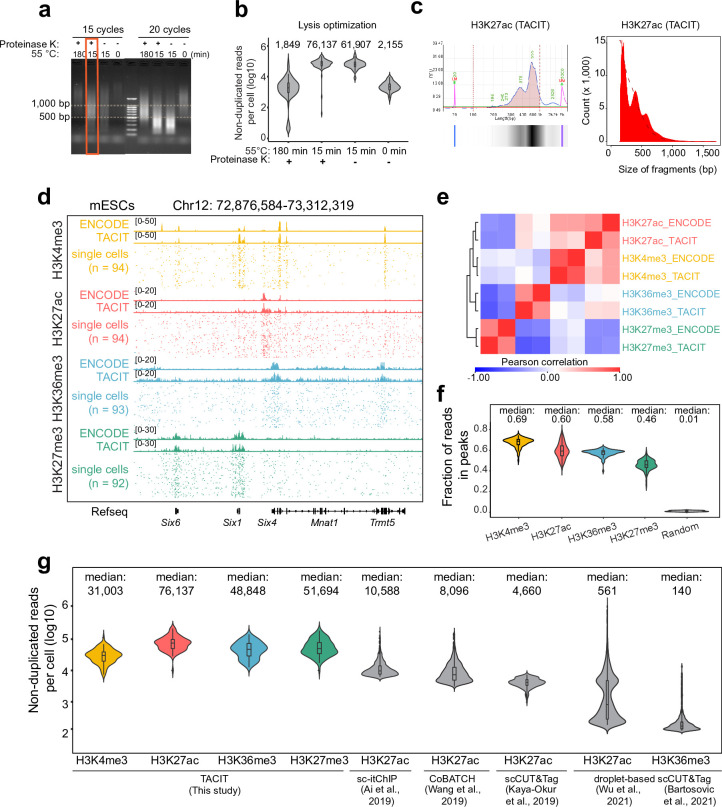
Substantial epigenetic resetting during early embryo development from fertilization to blastocyst formation ensures zygotic genome activation and leads to progressive cellular heterogeneities. Mapping single-cell epigenomic profiles of core histone modifications that cover each individual cell is a fundamental goal in developmental biology. Here we develop target chromatin indexing and tagmentation (TACIT), a method that enabled genome-coverage single-cell profiling of seven histone modifications across mouse early embryos. We integrated these single-cell histone modifications with single-cell RNA sequencing data to chart a single-cell resolution epigenetic landscape. Multimodal chromatin-state annotations showed that the onset of zygotic genome activation at the early two-cell stage already primes heterogeneities in totipotency. We used machine learning to identify totipotency gene regulatory networks, including stage-specific transposable elements and putative transcription factors. CRISPR activation of a combination of these identified transcription factors induced totipotency activation in mouse embryonic stem cells. Together with single-cell co-profiles of multiple histone modifications, we developed a model that predicts the earliest cell branching towards the inner cell mass and the trophectoderm in latent multimodal space and identifies regulatory elements and previously unknown lineage-specifying transcription factors. Our work provides insights into single-cell epigenetic reprogramming, multimodal regulation of cellular lineages and cell-fate priming during mouse pre-implantation development.
从受精到囊胚形成的早期胚胎发育过程中,大量的表观遗传重编程确保了合子基因组的激活,并导致细胞异质性的逐步增加。绘制覆盖每个细胞的核心组蛋白修饰的单细胞表观基因组图谱是发育生物学的一个基本目标。在这里,我们开发了靶向染色质索引和转座酶标签化技术(TACIT),这是一种能够对小鼠早期胚胎中的七种组蛋白修饰进行全基因组覆盖单细胞分析的方法。我们将这些单细胞组蛋白修饰与单细胞RNA测序数据整合起来,绘制了单细胞分辨率的表观遗传图谱。多模态染色质状态注释表明,在早期二细胞阶段合子基因组激活的开始已经引发了全能性的异质性。我们使用机器学习来识别全能性基因调控网络,包括阶段特异性转座元件和假定的转录因子。对这些鉴定出的转录因子进行组合的CRISPR激活可诱导小鼠胚胎干细胞中的全能性激活。结合多种组蛋白修饰的单细胞共分析,我们开发了一个模型,该模型可以预测在潜在多模态空间中最早向内细胞团和滋养外胚层分支的细胞,并识别调控元件和以前未知的谱系特异性转录因子。我们的工作为小鼠植入前发育过程中的单细胞表观遗传重编程、细胞谱系的多模态调控和细胞命运启动提供了见解。