Henry Y, Wood H, Morrissey J P, Petfalski E, Kearsey S, Tollervey D
EMBL, Heidelberg, Germany.
EMBO J. 1994 May 15;13(10):2452-63. doi: 10.1002/j.1460-2075.1994.tb06530.x.
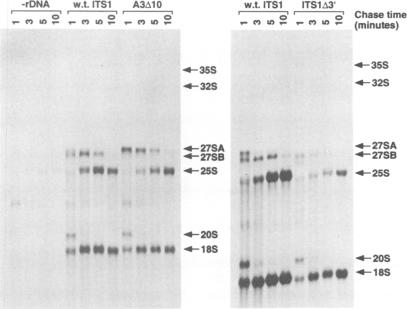
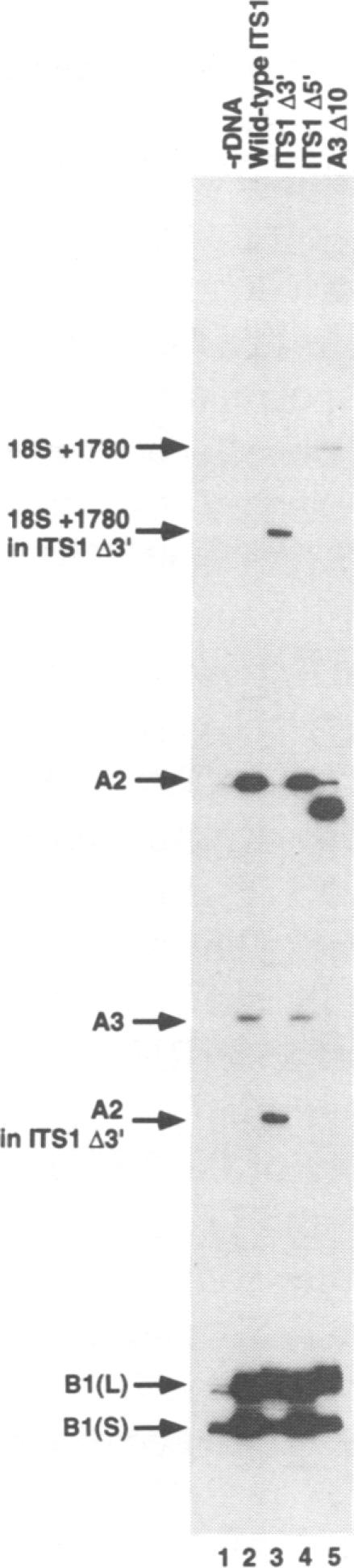
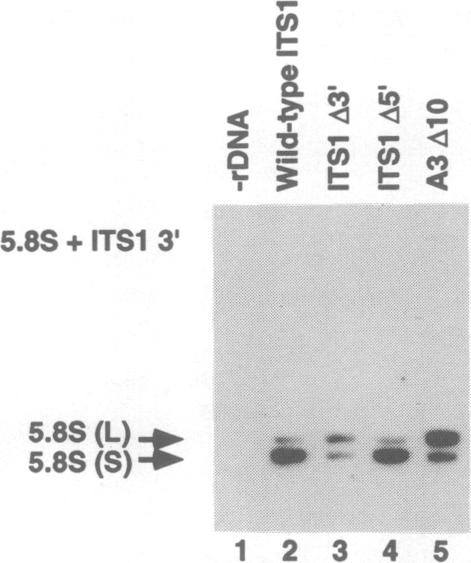
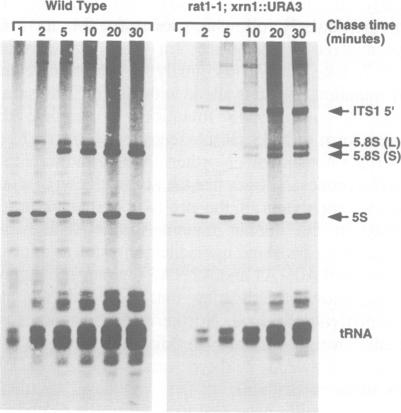
We have developed techniques for the detailed analysis of cis-acting sequences in the pre-rRNA of Saccharomyces cerevisiae and used these to study the processing of internal transcribed spacer 1 (ITS1) leading to the synthesis of 5.8S rRNA. As is the case for many eukaryotes, the 5' end of yeast 5.8S rRNA is heterogeneous; we designate the major, short form 5.8S(S), and the minor form (which is seven or eight nucleotides longer) 5.8S(L). These RNAs do not have a precursor/product relationship, but result from the use of alternative processing pathways. In the major pathway, a previously unidentified processing site in ITS1, designated A3, is cleaved. A 10 nucleotide deletion at site A3 strongly inhibits processing of A3 and the synthesis of 5.8S(S); processing is predominantly transferred to the alternative 5.8S(L) pathway. Site A3 lies 76 nucleotides 5' to the end of 5.8S(S), and acts as an entry site for 5'-->3' exonuclease digestion which generates the 5' end of 5.8S(S). This pathway is inhibited in strains mutant for XRN1p and RAT1p. Both of these proteins have been reported to have 5'-->3' exonuclease activity in vitro. Formation of 5.8S(L) is increased by mutations at A3 in cis or in RAT1p and XRN1p in trans, and is kinetically faster than 5.8S(S) synthesis.
我们已经开发出了用于详细分析酿酒酵母前体核糖体RNA(pre-rRNA)中顺式作用序列的技术,并利用这些技术来研究内部转录间隔区1(ITS1)的加工过程,该过程导致了5.8S核糖体RNA(rRNA)的合成。与许多真核生物的情况一样,酵母5.8S rRNA的5'端是异质的;我们将主要的短形式命名为5.8S(S),次要形式(比主要形式长7或8个核苷酸)命名为5.8S(L)。这些RNA不存在前体/产物关系,而是由不同的加工途径产生的。在主要途径中,ITS1中一个先前未鉴定的加工位点(命名为A3)被切割。A3位点的10个核苷酸缺失强烈抑制A3的加工和5.8S(S)的合成;加工过程主要转移到替代的5.8S(L)途径。A3位点位于5.8S(S)末端的5'端76个核苷酸处,并作为5'→3'核酸外切酶消化的入口位点,该消化产生5.8S(S)的5'端。在XRN1p和RAT1p突变的菌株中,该途径受到抑制。据报道,这两种蛋白质在体外都具有5'→3'核酸外切酶活性。顺式A3位点或反式RAT1p和XRN1p中的突变会增加5.8S(L)的形成,并且其动力学速度比5.8S(S)的合成更快。