Oh B K, Pace N R
Department of Chemistry, Indiana University, Bloomington 47405.
Nucleic Acids Res. 1994 Oct 11;22(20):4087-94. doi: 10.1093/nar/22.20.4087.
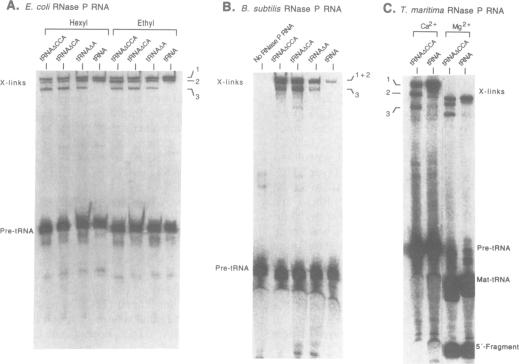
Ribonuclease P, which contains a catalytic RNA subunit, cleaves 5' precursor-specific sequences from pre-tRNAs. It was previously shown that the RNase P RNA optimally cleaves substrates which contain the mature, 3'-terminal CCA of tRNA. In order to determine the contributions of those individual 3'-terminal nucleotides to the interaction, pre-tRNAs that have CCA, only CC or C or are without CCA at the 3'-end were synthesized by run-off transcription, tested as substrates for cleavage by RNase P RNA and used in photoaffinity crosslinking experiments to examine contact sites in the ribozyme. In order to generalize the results, analyses were carried out using three different bacterial RNase P RNAs, from Escherichia coli, Bacillus subtilis and Thermotoga maritima. At optimal (Kcat/Km) ionic strength (1 M NH4+/25 mM Mg2+), Km increases incrementally 3- to 10-fold upon stepwise removal of each nucleotide from the 3'-end. At high ionic strength (2 M NH4+/50 mM Mg2+), which suppresses conformational effects, removal of the 3'-terminal A had little effect on Km, indicating that it is not a specific contact. Analysis of the deletion and substitution mutants indicated that the C residues act specially; their contribution to binding energy at high ionic strength (approximately 1 kcal/mol) is consistent with a non-Watson-Crick interaction, possibly irregular triple-strand formation with some component of the RNase P RNA. In agreement with previous studies, we find that the RNase P holoenzyme in vitro does not discriminate between tRNAs containing or lacking CCA. The structural elements of the three RNase P RNAs in proximity to the 3'-end of tRNA were examined by photoaffinity crosslinking. Photoagent-labeled tRNAs with 3'-terminal CCA, only CC or C, or lacking all these nucleotides were covalently conjugated to the three RNase P RNAs by irradiation and the sites of crosslinks were mapped by primer extension. The main crosslink sites are located in a highly conserved loop (probably an irregular helix) that is part of the core of the RNase P RNA secondary structure. The crosslinking results orient the CCA of tRNA with respect to that region of the RNase P RNA.
核糖核酸酶P含有一个催化性RNA亚基,可从前体tRNA上切割5'前体特异性序列。先前的研究表明,核糖核酸酶P RNA能最佳地切割含有成熟tRNA 3'-末端CCA的底物。为了确定这些单个3'-末端核苷酸对相互作用的贡献,通过连续转录合成了3'-末端具有CCA、仅含CC或C或无CCA的前体tRNA,将其作为核糖核酸酶P RNA切割的底物进行测试,并用于光亲和交联实验以检测核酶中的接触位点。为了使结果具有普遍性,使用了来自大肠杆菌、枯草芽孢杆菌和嗜热栖热菌的三种不同细菌核糖核酸酶P RNA进行分析。在最佳(Kcat/Km)离子强度(1 M NH4+/25 mM Mg2+)下,从3'-末端逐步去除每个核苷酸时,Km会逐渐增加3至10倍。在高离子强度(2 M NH4+/50 mM Mg2+)下,这种离子强度抑制了构象效应,去除3'-末端的A对Km影响很小,表明它不是特异接触。对缺失和替代突变体的分析表明,C残基具有特殊作用;它们在高离子强度下对结合能的贡献(约1千卡/摩尔)与非沃森-克里克相互作用一致,可能是与核糖核酸酶P RNA的某些成分形成不规则三链结构。与先前的研究一致,我们发现体外核糖核酸酶P全酶对含有或缺乏CCA的tRNA没有区分作用。通过光亲和交联研究了三种核糖核酸酶P RNA靠近tRNA 3'-末端的结构元件。用3'-末端具有CCA、仅含CC或C或缺乏所有这些核苷酸的光试剂标记的tRNA,通过照射与三种核糖核酸酶P RNA共价结合,并通过引物延伸确定交联位点。主要交联位点位于一个高度保守的环(可能是不规则螺旋)中,该环是核糖核酸酶P RNA二级结构核心的一部分。交联结果确定了tRNA的CCA相对于核糖核酸酶P RNA该区域的方向。