Colgan J, Manley J L
Department of Biological Sciences, Columbia University, New York, NY 10027.
Proc Natl Acad Sci U S A. 1995 Mar 14;92(6):1955-9. doi: 10.1073/pnas.92.6.1955.
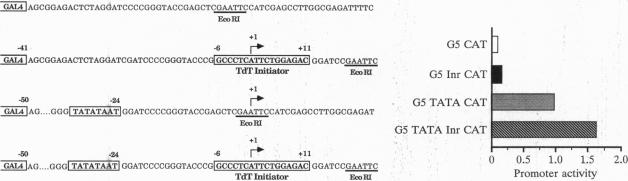
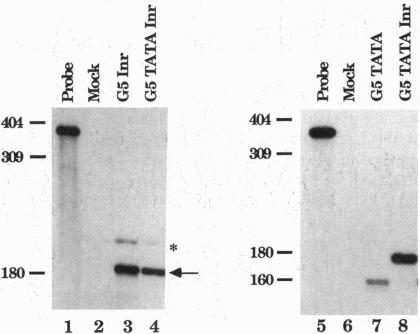
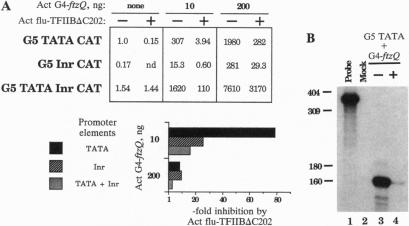
Core promoters for RNA polymerase II frequently contain either (or both) of two consensus sequence elements, a TATA box and/or an initiator (Inr). Using test promoters consisting of prototypical TATA and/or Inr elements, together with binding sites for sequence-specific activators, we have analyzed the function of TATA and Inr elements in vivo. In the absence of activators, the TATA element was significantly more active than the Inr, and the combination of elements was only slightly more effective than the TATA-only promoter. In the presence of any of several coexpressed activator proteins, the TATA elements was again most active, but here addition of the Inr allowed significant increases in activity, indicating a cooperative interaction between the two elements. An interesting exception was observed with the activator Sp1, which was more effective with the Inr-only promoter, and addition of a TATA box did not enhance activity. Finally, in all cases the TATA plus Inr promoters were found to be partially or completely resistant to the dominant negative effects of a transcription factor TFIIB mutant previously shown to interfere with expression from TATA-only promoters. This result strengthens the conclusion that TATA and Inr elements can cooperate in vivo.
RNA聚合酶II的核心启动子通常包含两个共有序列元件中的一个(或两个),即TATA盒和/或起始子(Inr)。我们使用由典型的TATA和/或Inr元件以及序列特异性激活因子的结合位点组成的测试启动子,在体内分析了TATA和Inr元件的功能。在没有激活因子的情况下,TATA元件比Inr元件活性显著更高,并且元件组合仅比仅含TATA的启动子略有效。在几种共表达的激活蛋白存在的情况下,TATA元件再次活性最高,但此时添加Inr可使活性显著增加,表明这两个元件之间存在协同相互作用。观察到一个有趣的例外情况,激活因子Sp1对仅含Inr的启动子更有效,添加TATA盒并未增强活性。最后,在所有情况下,发现TATA加Inr的启动子对先前显示会干扰仅含TATA启动子表达的转录因子TFIIB突变体的显性负效应具有部分或完全抗性。这一结果强化了TATA和Inr元件在体内可协同作用的结论。