Erickson H P
Department of Cell Biology, Duke University Medical Center, Durham, NC 27705.
Proc Natl Acad Sci U S A. 1994 Oct 11;91(21):10114-8. doi: 10.1073/pnas.91.21.10114.
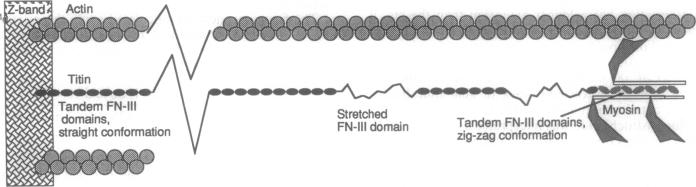
The elastic protein titin comprises a tandem array of fibronectin type III and immunoglobulin domains, which are structurally similar 7-strand beta-sandwiches. A proposed mechanism for stretching titin, by sequential denaturation of individual fibronectin type III-immunoglobulin domains in response to applied tension, is analyzed here quantitatively. The folded domain is approximately 4 nm long, and the unraveled polypeptide can extend to 29 nm, providing a 7-fold stretch over the relaxed length. Elastic recoil is achieved by refolding of the denatured domains when the force is released. The critical force required to denature a domain is calculated to be 3.5-5 pN, based on a net free energy for denaturation of 7-14 kcal/mol, plus 5 kcal/mol to extend the polypeptide (1 cal = 4.184 J). This force is comparable to the 2- to 7-pN force generated by single myosin or kinesin molecules. The force needed to pull apart a noncovalent protein-protein interface is estimated here to be 10-30 pN, implying that titin will stretch internally before the molecule is pulled from its attachment at the Z band. Many extracellular matrix and cell adhesion molecules, such as fibronectin, contain tandem arrays of fibronectin type III domains. Both single molecules and matrix fibers should have elastic properties similar to titin.
弹性蛋白肌联蛋白由串联排列的纤连蛋白III型和免疫球蛋白结构域组成,这些结构域在结构上类似7股β折叠三明治。本文定量分析了一种关于肌联蛋白拉伸的推测机制,即单个纤连蛋白III型-免疫球蛋白结构域响应施加的张力而依次变性。折叠后的结构域约4纳米长,解折叠的多肽可延伸至29纳米,相比松弛长度有7倍的拉伸。当力释放时,变性结构域重新折叠实现弹性回缩。基于变性的净自由能为7-14千卡/摩尔,再加上5千卡/摩尔用于多肽延伸(1卡 = 4.184焦耳),计算得出使一个结构域变性所需的临界力为3.5-5皮牛。该力与单个肌球蛋白或驱动蛋白分子产生的2-7皮牛的力相当。此处估计拉开非共价蛋白质-蛋白质界面所需的力为10-30皮牛,这意味着在分子从其在Z带的附着点被拉开之前,肌联蛋白会先在内部伸展。许多细胞外基质和细胞粘附分子,如纤连蛋白,都含有纤连蛋白III型结构域的串联阵列。单分子和基质纤维都应具有与肌联蛋白相似的弹性特性。