Tomkinson B, Robertson E, Kieff E
Department of Microbiology and Molecular Genetics and Medicine, Harvard University, Boston, Massachusetts 02115.
J Virol. 1993 Apr;67(4):2014-25. doi: 10.1128/JVI.67.4.2014-2025.1993.
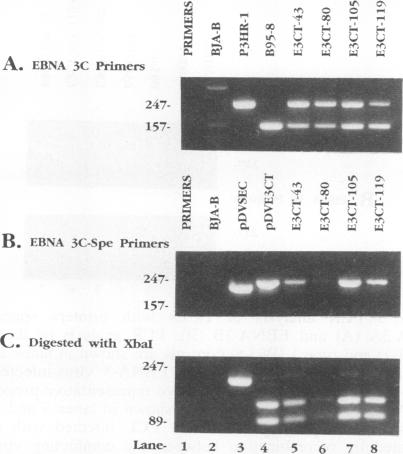
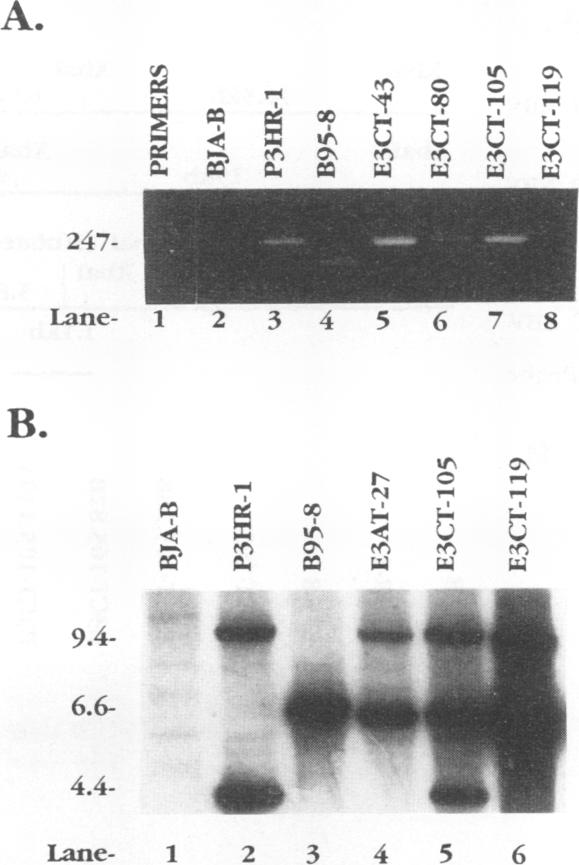
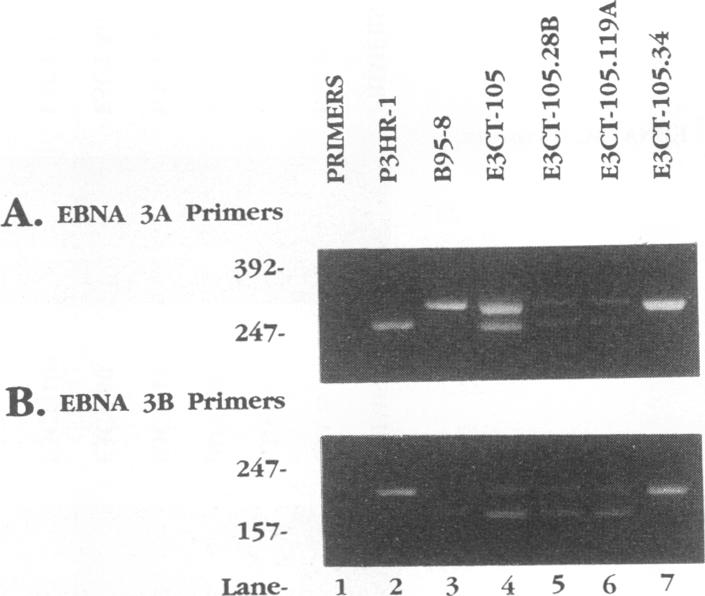
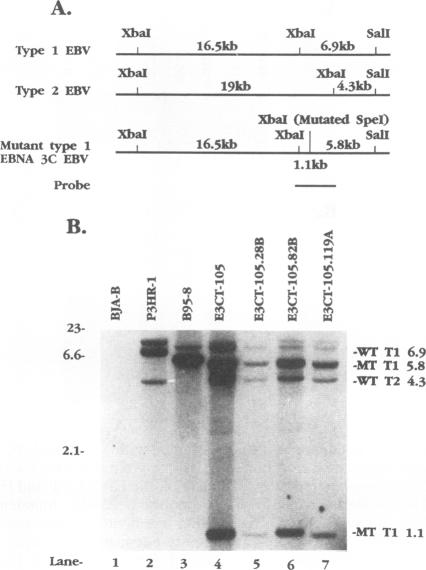
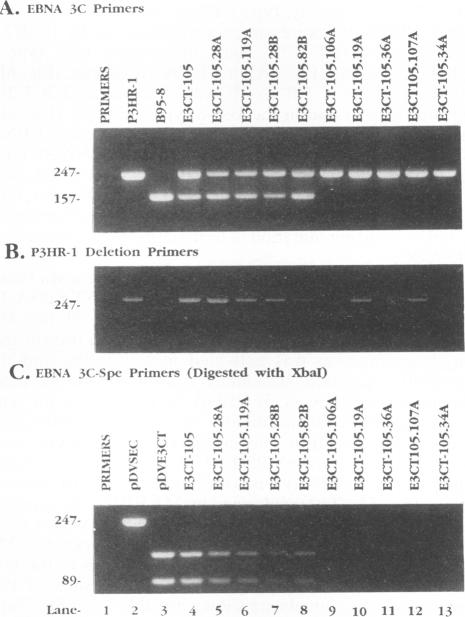
Recombinant Epstein-Barr viruses (EBV) with a translation termination codon mutation inserted into the nuclear protein 3A (EBNA-3A) or 3C (EBNA-3C) open reading frame were generated by second-site homologous recombination. These mutant viruses were used to infect primary B lymphocytes to assess the requirement of EBNA-3A or -3C for growth transformation. The frequency of obtaining transformants infected with a wild-type EBNA-3A recombinant EBV was 10 to 15%. In contrast, the frequency of obtaining transformants infected with a mutant EBNA-3A recombinant EBV was only 1.4% (9 mutants in 627 transformants analyzed). Transformants infected with mutant EBNA-3A recombinant virus could be obtained only by coinfection with another transformation-defective EBV which provided wild-type EBNA-3A in trans. Cells infected with mutant EBNA-3A recombinant virus lost the EBNA-3A mutation with expansion of the culture. The decreased frequency of recovery of the EBNA-3A mutation, the requirement for transformation-defective EBV coinfection, and the inability to maintain the EBNA-3A mutation indicate that EBNA-3A is essential or critical for lymphocyte growth transformation and that the EBNA-3A mutation has a partial dominant negative effect. Five transformants infected with mutant EBNA-3C recombinant virus EBV were also identified and expanded. All five also required wild-type EBNA-3C in trans. Serial passage of the mutant recombinant virus into primary B lymphocytes resulted in transformants only when wild-type EBNA-3C was provided in trans by coinfection with a transformation-defective EBV carrying a wild-type EBNA-3C gene. A secondary recombinant virus in which the mutated EBNA-3C gene was replaced by wild-type EBNA-3C was able to transform B lymphocytes. Thus, EBNA-3C is also essential or critical for primary B-lymphocyte growth transformation.
通过第二位点同源重组产生了在核蛋白3A(EBNA - 3A)或3C(EBNA - 3C)开放阅读框中插入翻译终止密码子突变的重组爱泼斯坦 - 巴尔病毒(EBV)。这些突变病毒用于感染原代B淋巴细胞,以评估EBNA - 3A或 - 3C对生长转化的需求。获得感染野生型EBNA - 3A重组EBV的转化体的频率为10%至15%。相比之下,获得感染突变型EBNA - 3A重组EBV的转化体的频率仅为1.4%(在分析的627个转化体中有9个突变体)。仅通过与另一种提供野生型EBNA - 3A反式互补的转化缺陷型EBV共感染,才能获得感染突变型EBNA - 3A重组病毒的转化体。随着培养物的扩增,感染突变型EBNA - 3A重组病毒的细胞失去了EBNA - 3A突变。EBNA - 3A突变恢复频率的降低、对转化缺陷型EBV共感染的需求以及无法维持EBNA - 3A突变表明,EBNA - 3A对淋巴细胞生长转化至关重要,并且EBNA - 3A突变具有部分显性负效应。还鉴定并扩增了5个感染突变型EBNA - 3C重组病毒EBV的转化体。所有5个转化体也都需要野生型EBNA - 3C的反式互补。当通过与携带野生型EBNA - 3C基因的转化缺陷型EBV共感染提供野生型EBNA - 3C反式互补时,突变重组病毒连续传代至原代B淋巴细胞才能产生转化体。一种二级重组病毒,其中突变的EBNA - 3C基因被野生型EBNA - 3C取代,能够转化B淋巴细胞。因此,EBNA - 3C对原代B淋巴细胞生长转化也至关重要。