Ludewig U, Jentsch T J, Pusch M
Center for Molecular Neurobiology (ZMNH), Hamburg University, D-20246 Hamburg, Germany.
J Gen Physiol. 1997 Aug;110(2):165-71. doi: 10.1085/jgp.110.2.165.
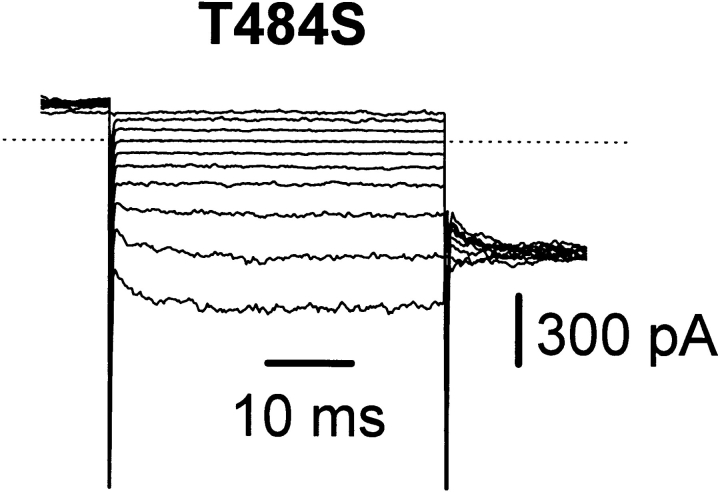
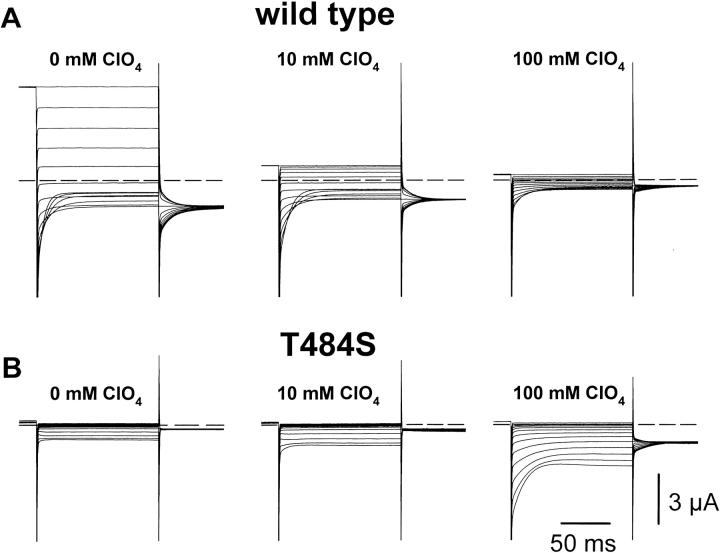
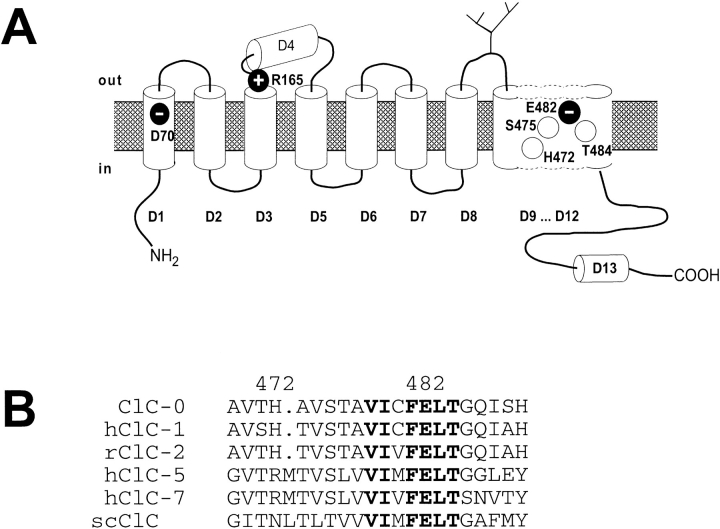
Several cloned ClC-type Cl- channels open and close in a voltage-dependent manner. The Torpedo electric organ Cl- channel, ClC-0, is the best studied member of this gene family. ClC-0 is gated by a fast and a slow gating mechanism of opposite voltage direction. Fast gating is dependent on voltage and on the external and internal Cl- concentration, and it has been proposed that the permeant anion serves as the gating charge in ClC-0 (Pusch, M., U. Ludewig, A. Rehfeldt, and T.J. Jentsch. 1995. Nature (Lond.). 373:527-531). The deactivation at negative voltages of the muscular ClC-1 channel is similar but not identical to ClC-0. Different from the extrinsic voltage dependence suggested for ClC-0, an intrinsic voltage sensor had been proposed to underlie the voltage dependence in ClC-1 (Fahlke, C., R. Rüdel, N. Mitrovic, M. Zhou, and A.L. George. 1995. Neuron. 15:463-472; Fahlke, C., A. Rosenbohm, N. Mitrovic, A.L. George, and R. Rüdel. 1996. Biophys. J. 71:695-706). The gating model for ClC-1 was partially based on the properties of a point-mutation found in recessice myotonia (D136G). Here we investigate the functional effects of mutating the corresponding residue in ClC-0 (D70). Both the corresponding charge neutralization (D70G) and a charge conserving mutation (D70E) led to an inwardly rectifying phenotype resembling that of ClC-1 (D136G). Several other mutations at very different positions in ClC-0 (K165R, H472K, S475T, E482D, T484S, T484Q), however, also led to a similar phenotype. In one of these mutants (T484S) the typical wild-type gating, characterized by a deactivation at negative voltages, can be partially restored by using external perchlorate (ClO4-) solutions. We conclude that gating in ClC-0 and ClC-1 is due to similar mechanisms. The negative charge at position 70 in ClC-0 does not specifically confer the voltage sensitivity in ClC-channels, and there is no need to postulate an intrinsic voltage sensor in ClC-channels.
几种克隆的ClC型Cl⁻通道以电压依赖的方式打开和关闭。电鳐电器官的Cl⁻通道ClC-0是该基因家族中研究得最透彻的成员。ClC-0由电压方向相反的快速和慢速门控机制控制。快速门控依赖于电压以及细胞外和细胞内的Cl⁻浓度,有人提出通透阴离子在ClC-0中充当门控电荷(普施,M.,U. 路德维希,A. 雷费尔特,和T.J. 延奇。1995年。《自然》(伦敦)。373:527 - 531)。肌肉ClC-1通道在负电压下的失活与ClC-0相似但不完全相同。与ClC-0所暗示的外在电压依赖性不同,有人提出ClC-1中的电压依赖性是由一个内在电压传感器引起的(法尔克,C.,R. 吕德尔,N. 米特罗维奇,M. 周,和A.L. 乔治。1995年。《神经元》。15:463 - 472;法尔克,C.,A. 罗森博姆,N. 米特罗维奇,A.L. 乔治,和R. 吕德尔。1996年。《生物物理学杂志》。71:695 - 706)。ClC-1的门控模型部分基于在隐性肌强直中发现的一个点突变(D136G)的特性。在这里,我们研究了ClC-0中相应残基(D70)突变的功能效应。相应的电荷中和(D70G)和一个电荷保守突变(D70E)都导致了类似于ClC-1(D136G)的内向整流表型。然而,ClC-0中其他几个位于非常不同位置的突变(K165R、H472K、S475T、E482D、T484S、T484Q)也导致了类似的表型。在其中一个突变体(T484S)中,以负电压下失活为特征的典型野生型门控可以通过使用外部高氯酸盐(ClO₄⁻)溶液部分恢复。我们得出结论,ClC-0和ClC-1中的门控是由相似的机制引起的。ClC-0中70位的负电荷并非专门赋予ClC通道电压敏感性,并且无需假定ClC通道中有内在电压传感器。