Ugulava Natalia B, Frederick Kendra K, Jarrett Joseph T
Johnson Research Foundation and Department of Biochemistry and Biophysics, University of Pennsylvania, Philadelphia, Pennsylvania 19104-6059, USA.
Biochemistry. 2003 Mar 11;42(9):2708-19. doi: 10.1021/bi0261084.
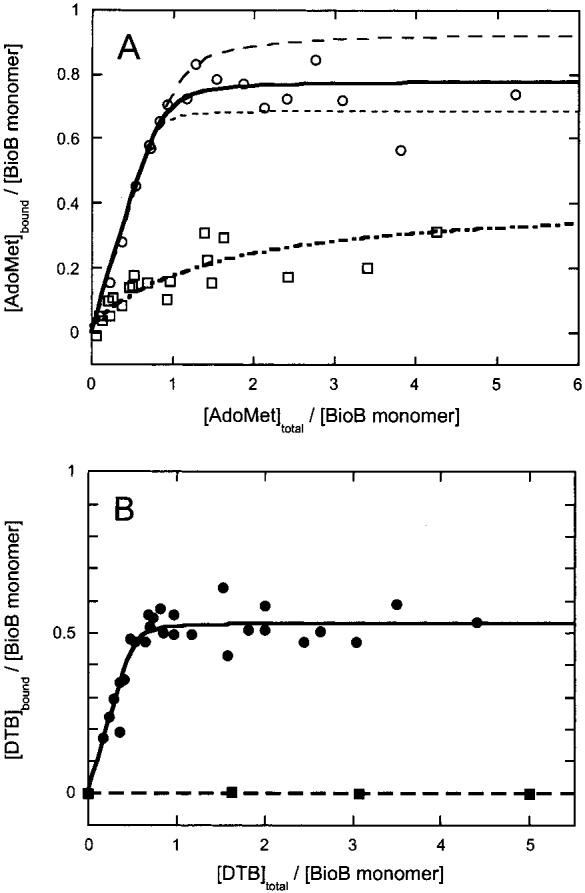
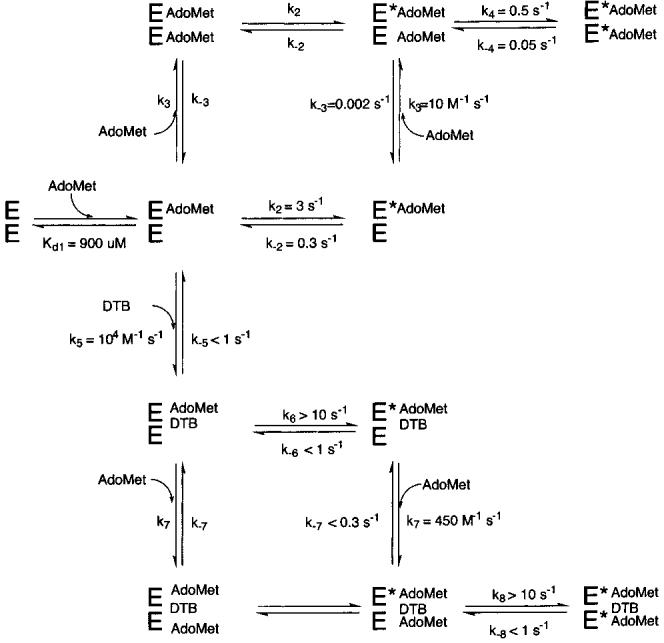
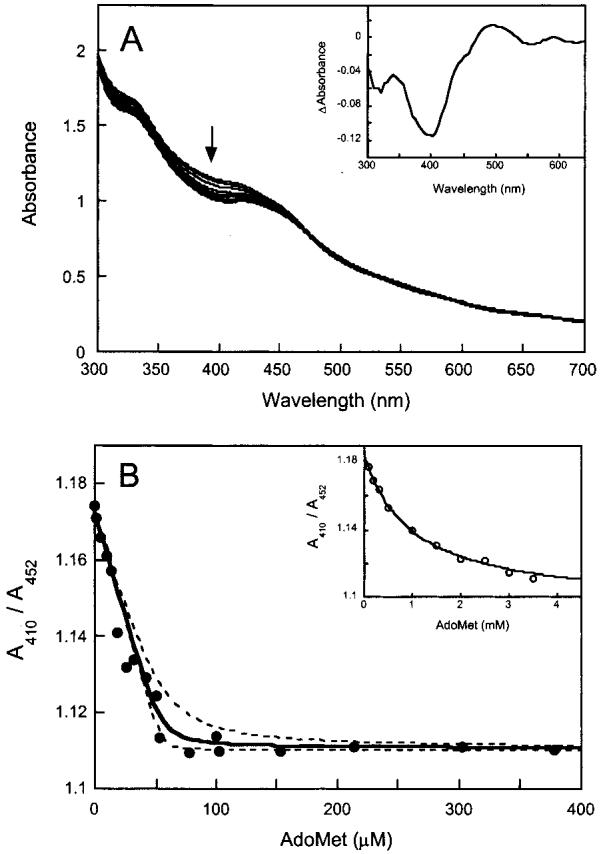
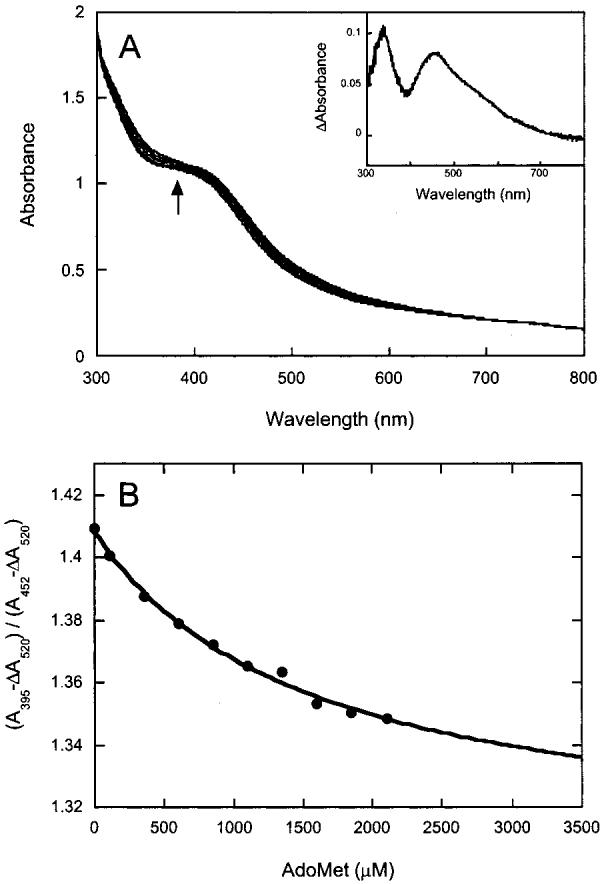
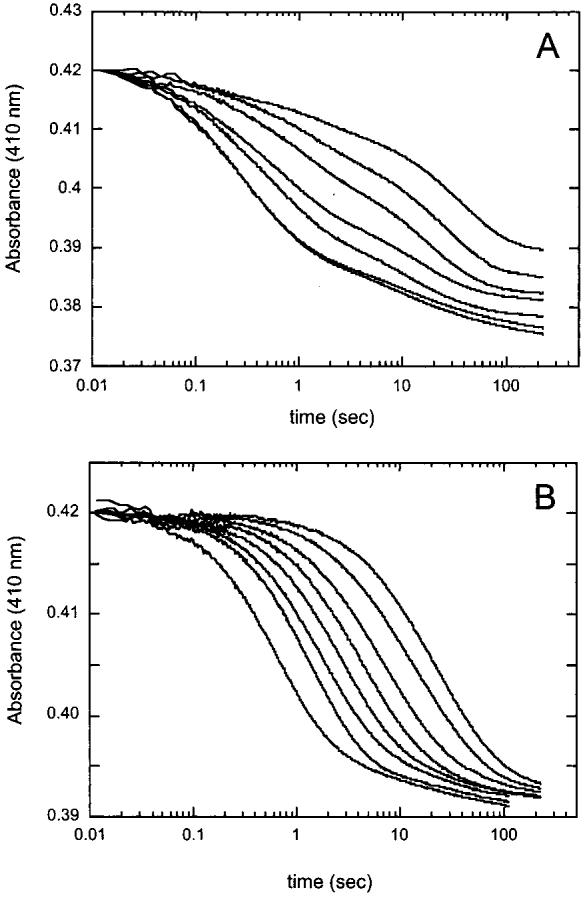
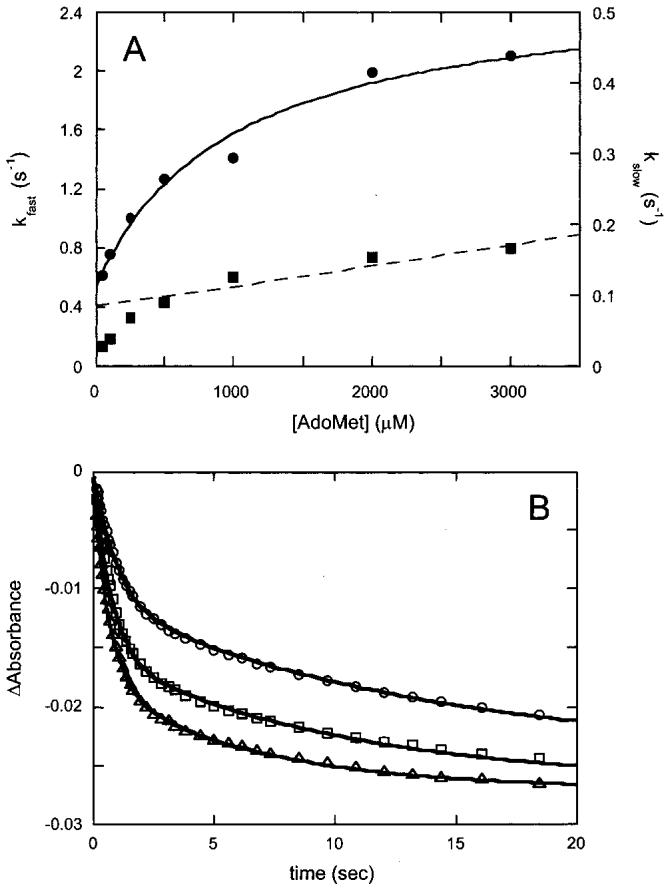
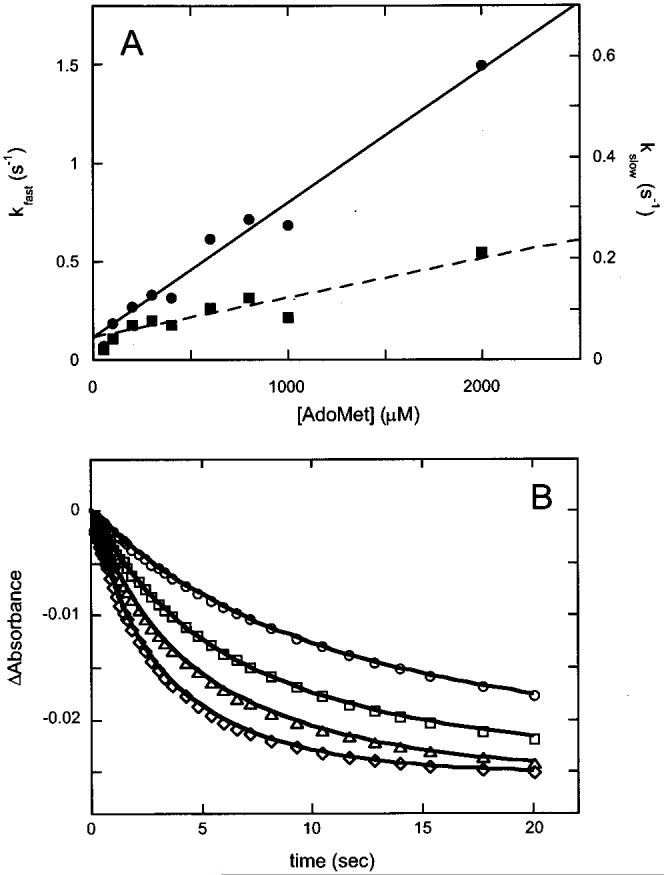
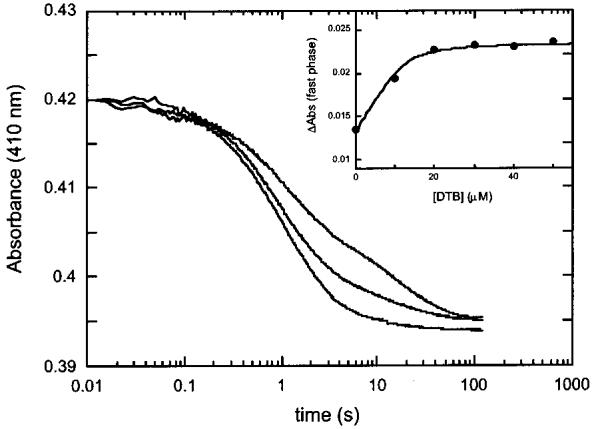
Biotin synthase (BS) is an AdoMet-dependent radical enzyme that catalyzes the insertion of sulfur into saturated C6 and C9 atoms in the substrate dethiobiotin. To facilitate sulfur insertion, BS catalyzes the reductive cleavage of AdoMet to methionine and 5'-deoxyadenosyl radicals, which then abstract hydrogen atoms from the C6 and C9 positions of dethiobiotin. The enzyme from Escherichia coli is purified as a dimer that contains one [2Fe-2S]2+ cluster per monomer and can be reconstituted in vitro to contain an additional [4Fe-4S]2+ cluster per monomer. Since each monomer contains each type of cluster, the dimeric enzyme could contain one active site per monomer, or could contain a single active site at the dimer interface. To address these possibilities, and to better understand the manner in which biotin synthase controls radical generation and reactivity, we have examined the binding of AdoMet and DTB to reconstituted biotin synthase. We find that both the [2Fe-2S]2+ cluster and the [4Fe-4S]2+ cluster must be present for tight substrate binding. Further, substrate binding is highly cooperative, with the affinity for AdoMet increasing >20-fold in the presence of DTB, while DTB binds only in the presence of AdoMet. The stoichiometry of binding is ca. 2:1:1 AdoMet:DTB:BS dimer, suggesting that biotin synthase has a single functional active site per dimer. AdoMet binding, either in the presence or in the absence of DTB, leads to a decrease in the magnitude of the UV-visible absorption band at approximately 400 nm that we attribute to changes in the coordination environment of the [4Fe-4S]2+ cluster. Using these spectral changes as a probe, we have examined the kinetics of AdoMet and DTB binding, and propose an ordered binding mechanism that is followed by a conformational change in the enzyme-substrate complex. This kinetic analysis suggests that biotin synthase is evolved to bind AdoMet both weakly and slowly in the absence of DTB, while both the rate of binding and the affinity for AdoMet are increased in the presence of DTB. Cooperative binding of AdoMet and DTB may be an important mechanism for limiting the production of 5'-deoxyadenosyl radicals in the absence of the correct substrate.
生物素合酶(BS)是一种依赖于腺苷甲硫氨酸(AdoMet)的自由基酶,它催化将硫插入底物脱硫生物素中饱和的C6和C9原子上。为了促进硫的插入,BS催化AdoMet还原裂解为甲硫氨酸和5'-脱氧腺苷自由基,然后这些自由基从脱硫生物素的C6和C9位置夺取氢原子。来自大肠杆菌的这种酶以二聚体形式纯化,每个单体含有一个[2Fe-2S]2+簇,并且可以在体外重构为每个单体额外含有一个[4Fe-4S]2+簇。由于每个单体都含有每种类型的簇,二聚体酶可能每个单体含有一个活性位点,或者可能在二聚体界面处含有一个单一的活性位点。为了解决这些可能性,并更好地理解生物素合酶控制自由基产生和反应性的方式,我们研究了AdoMet和DTB与重构的生物素合酶的结合。我们发现,紧密的底物结合需要同时存在[2Fe-2S]2+簇和[4Fe-4S]2+簇。此外,底物结合具有高度协同性,在DTB存在的情况下,对AdoMet的亲和力增加超过20倍,而DTB仅在AdoMet存在时结合。结合的化学计量比约为2:1:1,即AdoMet:DTB:BS二聚体,这表明生物素合酶每个二聚体有一个单一的功能活性位点。无论是否存在DTB,AdoMet的结合都会导致约400nm处紫外可见吸收带的强度降低,我们将其归因于[4Fe-4S]2+簇配位环境的变化。利用这些光谱变化作为探针,我们研究了AdoMet和DTB结合的动力学,并提出了一种有序结合机制,随后酶-底物复合物会发生构象变化。这种动力学分析表明,生物素合酶在没有DTB的情况下进化为以较弱和较慢的方式结合AdoMet,而在DTB存在时,结合速率和对AdoMet的亲和力都会增加。AdoMet和DTB的协同结合可能是在没有正确底物时限制5'-脱氧腺苷自由基产生的一种重要机制。