Franceschi Renny T, Xiao Guozhi, Jiang Di, Gopalakrishnan Rajaram, Yang Shuying, Reith Elizabeth
Department of Periodontics/Prevention/Geriatrics, School of Medicine, University of Michigan, Ann Arbor, Michigan 48109-1078, USA.
Connect Tissue Res. 2003;44 Suppl 1(Suppl 1):109-16.
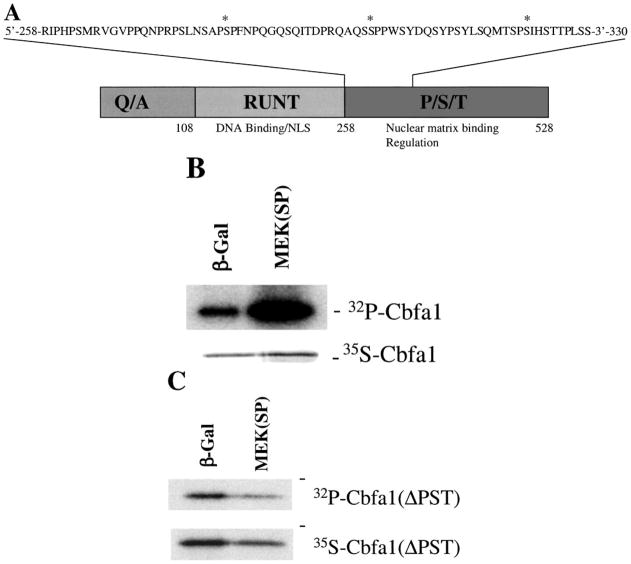
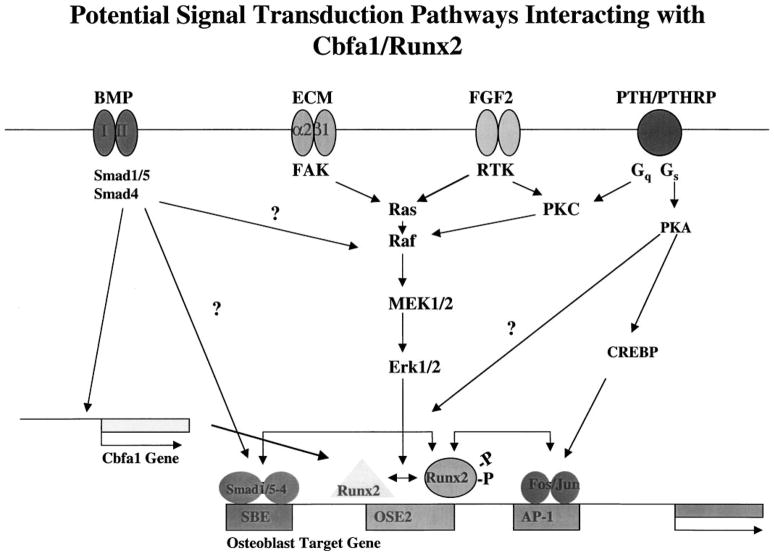
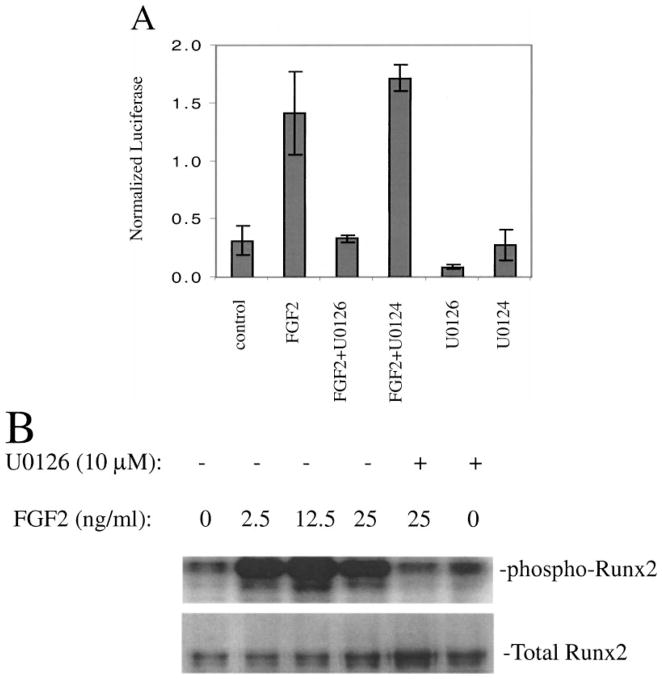
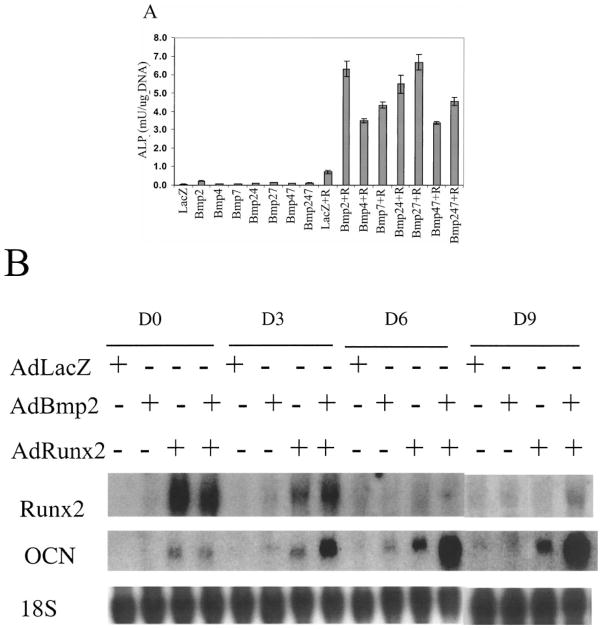
The Cbfa1/Runx2 transcription factor is essential for osteoblast differentiation. However, levels of Runx2 are often not well correlated with its transcriptional activity suggesting that this factor must be activated either by covalent modification or through interactions with other nuclear components. Runx2 is phosphorylated and activated by the mitogen-activated protein kinase (MAPK) pathway. This pathway is stimulated in at least two ways: by binding of type I collagen to alpha2beta1 integrins on the osteoblast surface and by treatment of cells with the osteogenic growth factor, FGF2. Protein kinase A (PKA) also may phosphorylate/activate Runx2 under certain conditions. Runx2 activity also is enhanced by factors known to stimulate specific signal transduction pathways such as PTH/PTHrP (signals through PKA and PKC pathways) and BMPs (Signal through Smad proteins). Interactions with Runx2 are complex involving both binding of distinct components such as AP-1 factors and Smads to separate sites on DNA, direct interactions between Runx2 and AP-1/Smad factors and MAPK or PKA-dependent Runx2 phosphorylation. These findings suggest that Runx2 plays a central role in coordinating multiple signals involved in osteoblast differentiation.
Cbfa1/Runx2转录因子对成骨细胞分化至关重要。然而,Runx2的水平与其转录活性往往没有很好的相关性,这表明该因子必须通过共价修饰或与其他核成分相互作用才能被激活。Runx2可被丝裂原活化蛋白激酶(MAPK)途径磷酸化并激活。该途径至少通过两种方式被刺激:通过I型胶原与成骨细胞表面的α2β1整合素结合,以及用成骨生长因子FGF2处理细胞。蛋白激酶A(PKA)在某些条件下也可能使Runx2磷酸化/激活。已知刺激特定信号转导途径的因子,如甲状旁腺激素/甲状旁腺激素相关蛋白(通过PKA和PKC途径发出信号)和骨形态发生蛋白(通过Smad蛋白发出信号),也可增强Runx2的活性。与Runx2的相互作用很复杂,涉及不同成分(如AP-1因子和Smads)与DNA上不同位点的结合、Runx2与AP-1/Smad因子之间的直接相互作用以及MAPK或PKA依赖的Runx2磷酸化。这些发现表明,Runx2在协调成骨细胞分化中涉及的多种信号方面发挥着核心作用。