Tanida Isei, Ueno Takashi, Kominami Eiki
Department of Biochemistry, School of Medicine, Juntendo University, 2-1-1 Hongo, Bunkyo-ku, Tokyo 113-8421, Japan.
Int J Biochem Cell Biol. 2004 Dec;36(12):2503-18. doi: 10.1016/j.biocel.2004.05.009.
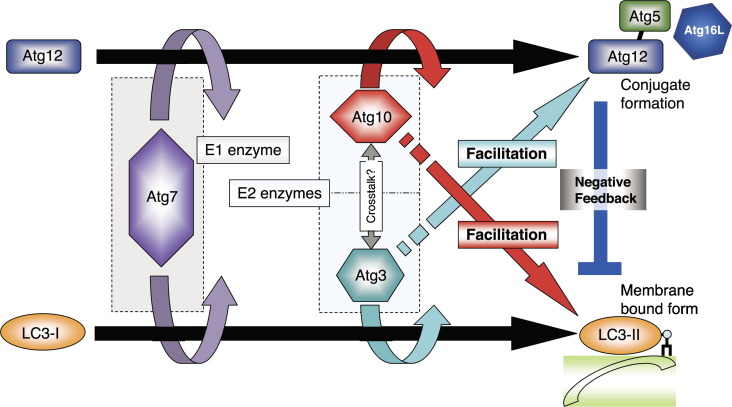
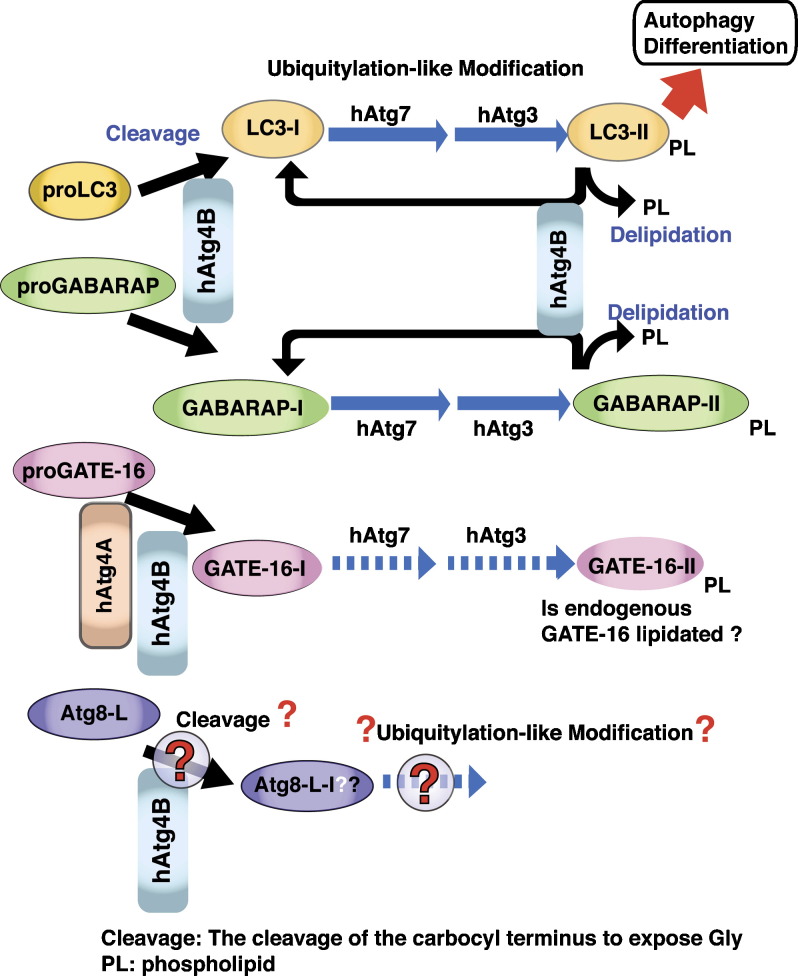
Autophagy is the bulk degradation of proteins and organelles, a process essential for cellular maintenance, cell viability, differentiation and development in mammals. Autophagy has significant associations with neurodegenerative diseases, cardiomyopathies, cancer, programmed cell death, and bacterial and viral infections. During autophagy, a cup-shaped structure, the preautophagosome, engulfs cytosolic components, including organelles, and closes, forming an autophagosome, which subsequently fuses with a lysosome, leading to the proteolytic degradation of internal components of the autophagosome by lysosomal lytic enzymes. During the formation of mammalian autophagosomes, two ubiquitylation-like modifications are required, Atg12-conjugation and LC3-modification. LC3 is an autophagosomal ortholog of yeast Atg8. A lipidated form of LC3, LC3-II, has been shown to be an autophagosomal marker in mammals, and has been used to study autophagy in neurodegenerative and neuromuscular diseases, tumorigenesis, and bacterial and viral infections. The other Atg8 homologues, GABARAP and GATE-16, are also modified by the same mechanism. In non-starved rats, the tissue distribution of LC3-II differs from those of the lipidated forms of GABARAP and GATE-16, GABARAP-II and GATE-16-II, suggesting that there is a functional divergence among these three modified proteins. Delipidation of LC3-II and GABARAP-II is mediated by hAtg4B. We review the molecular mechanism of LC3-modification, the crosstalk between LC3-modification and mammalian Atg12-conjugation, and the cycle of LC3-lipidation and delipidation mediated by hAtg4B, as well as recent findings concerning the other two Atg8 homologues, GABARAP and GATE-16. We also highlight recent findings regarding the pathobiology of LC3-modification, including its role in microbial infection, cancer and neuromuscular diseases.
自噬是蛋白质和细胞器的大量降解过程,这一过程对于哺乳动物的细胞维持、细胞活力、分化及发育至关重要。自噬与神经退行性疾病、心肌病、癌症、程序性细胞死亡以及细菌和病毒感染有着密切关联。在自噬过程中,一种杯状结构,即前自噬体,会吞噬包括细胞器在内的胞质成分,然后封闭形成自噬体,自噬体随后与溶酶体融合,导致自噬体内部成分被溶酶体裂解酶进行蛋白水解降解。在哺乳动物自噬体形成过程中,需要两种类泛素化修饰,即Atg12缀合和LC3修饰。LC3是酵母Atg8的自噬体直系同源物。LC3的脂化形式LC3-II已被证明是哺乳动物中的自噬体标志物,并已用于研究神经退行性和神经肌肉疾病、肿瘤发生以及细菌和病毒感染中的自噬。其他Atg8同源物GABARAP和GATE-16也通过相同机制进行修饰。在未饥饿的大鼠中,LC3-II的组织分布与GABARAP和GATE-16的脂化形式GABARAP-II和GATE-16-II不同,这表明这三种修饰蛋白之间存在功能差异。LC3-II和GABARAP-II的去脂化由hAtg4B介导。我们综述了LC3修饰的分子机制、LC3修饰与哺乳动物Atg12缀合之间的相互作用、由hAtg4B介导的LC3脂化和去脂化循环,以及关于其他两种Atg8同源物GABARAP和GATE-16的最新研究发现。我们还重点介绍了关于LC3修饰病理生物学的最新研究发现,包括其在微生物感染、癌症和神经肌肉疾病中的作用。