MRC Laboratory for Molecular Cell Biology, University College London, London WC1E 6BT, United Kingdom.
MRC Laboratory for Molecular Cell Biology, University College London, London WC1E 6BT, United Kingdom
J Biol Chem. 2019 Aug 23;294(34):12610-12621. doi: 10.1074/jbc.AC119.009977. Epub 2019 Jul 17.
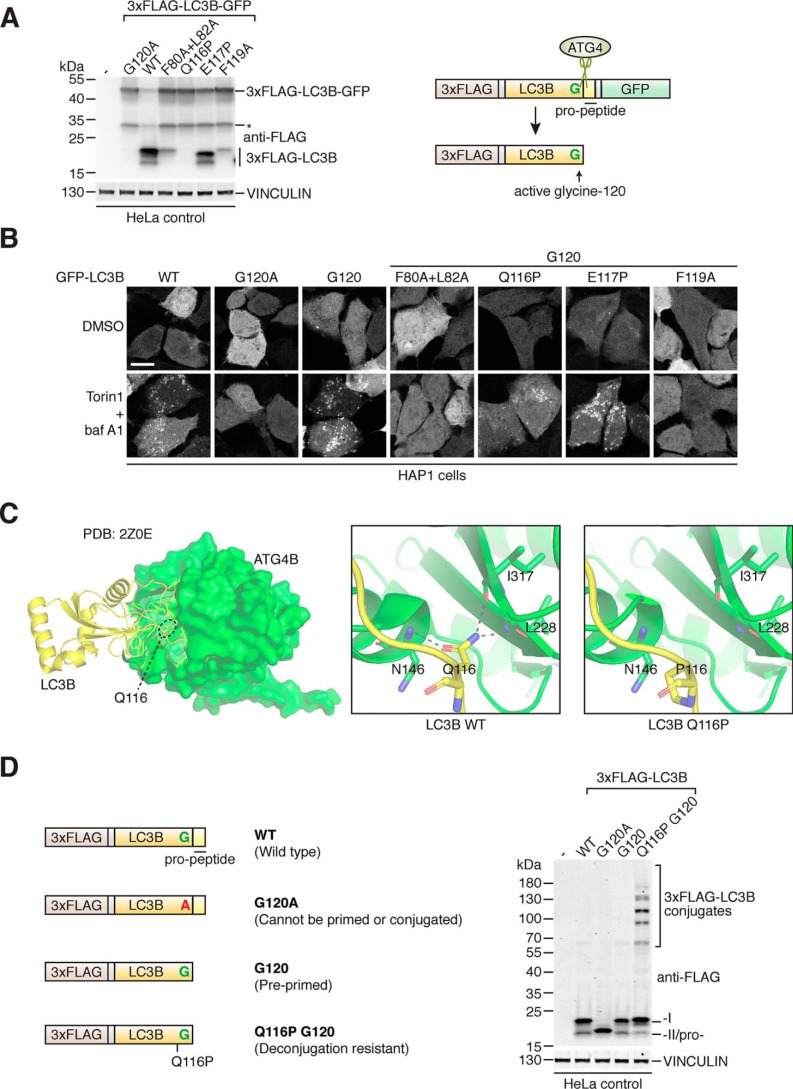
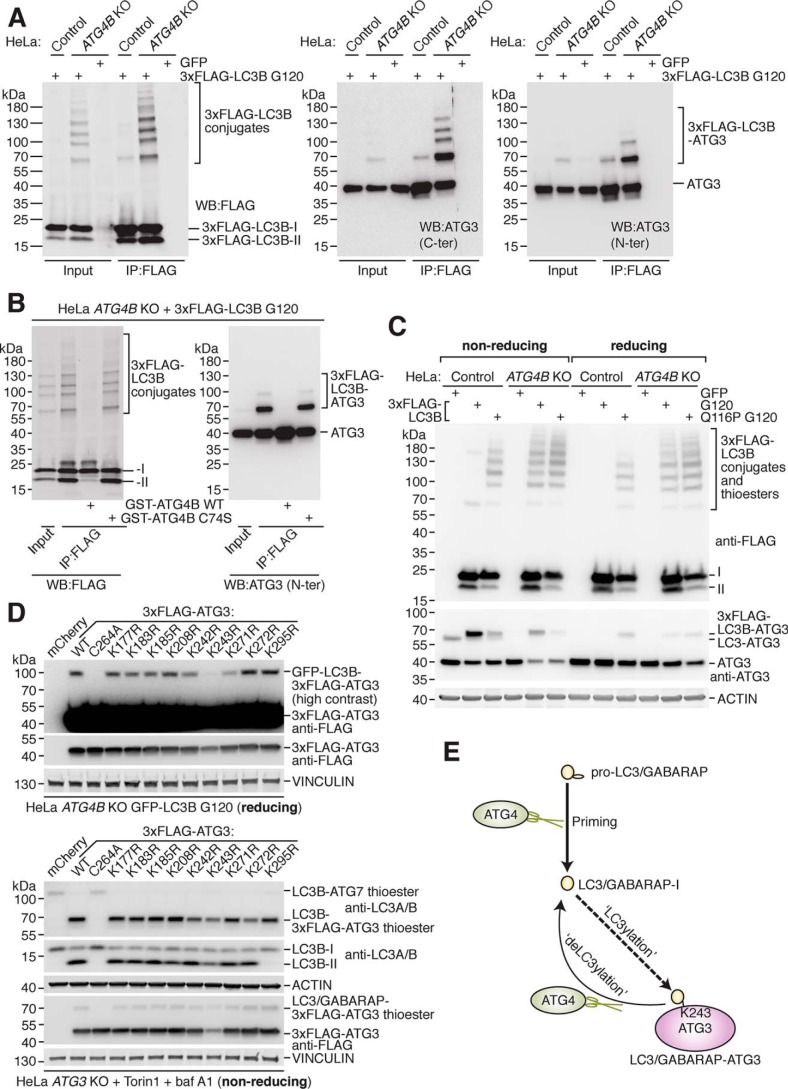
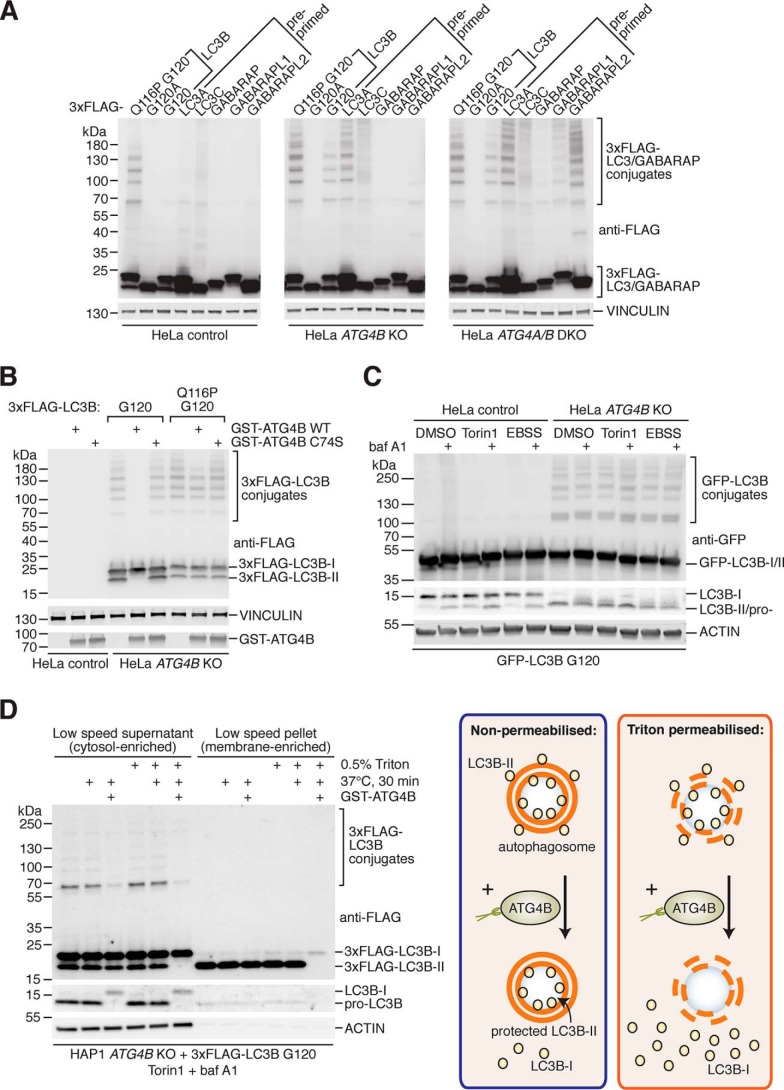
Microtubule-associated protein 1 light chain 3 α (LC3)/GABA type A receptor-associated protein (GABARAP) comprises a family of ubiquitin-like proteins involved in (macro)autophagy, an important intracellular degradation pathway that delivers cytoplasmic material to lysosomes via double-membrane vesicles called autophagosomes. The only currently known cellular molecules covalently modified by LC3/GABARAP are membrane phospholipids such as phosphatidylethanolamine in the autophagosome membrane. Autophagy-related 4 cysteine peptidase (ATG4) proteases process inactive pro-LC3/GABARAP before lipidation, and the same proteases can also deconjugate LC3/GABARAP from lipids. To determine whether LC3/GABARAP has other molecular targets, here we generated a pre-processed LC3B mutant (Q116P) that is resistant to ATG4-mediated deconjugation. Upon expression in human cells and when assessed by immunoblotting under reducing and denaturing conditions, deconjugation-resistant LC3B accumulated in multiple forms and at much higher molecular weights than free LC3B. We observed a similar accumulation when pre-processed versions of all mammalian LC3/GABARAP isoforms were expressed in ATG4-deficient cell lines, suggesting that LC3/GABARAP can attach also to other larger molecules. We identified ATG3, the E2-like enzyme involved in LC3/GABARAP lipidation, as one target of conjugation with multiple copies of LC3/GABARAP. We show that LC3B-ATG3 conjugates are distinct from the LC3B-ATG3 thioester intermediate formed before lipidation, and we biochemically demonstrate that ATG4B can cleave LC3B-ATG3 conjugates. Finally, we determined ATG3 residue Lys-243 as an LC3B modification site. Overall, we provide the first cellular evidence that mammalian LC3/GABARAP post-translationally modifies proteins akin to ubiquitination ("LC3ylation"), with ATG4 proteases acting like deubiquitinating enzymes to counteract this modification ("deLC3ylation").
微管相关蛋白 1 轻链 3α(LC3)/GABA 型 A 受体相关蛋白(GABARAP)属于一类泛素样蛋白,参与(巨)自噬,这是一种重要的细胞内降解途径,通过双层囊泡(自噬体)将细胞质物质递送至溶酶体。目前唯一已知的被 LC3/GABARAP 共价修饰的细胞内分子是质膜磷脂,如自噬体膜中的磷脂酰乙醇胺。自噬相关 4 半胱氨酸肽酶(ATG4)蛋白酶在脂质化之前处理非活性原 LC3/GABARAP,并且相同的蛋白酶也可以从脂质上脱离 LC3/GABARAP。为了确定 LC3/GABARAP 是否具有其他分子靶标,我们在此生成了一种预处理的 LC3B 突变体(Q116P),该突变体对 ATG4 介导的去共轭具有抗性。在人细胞中表达并在还原和变性条件下通过免疫印迹进行评估时,去共轭抗性 LC3B 以多种形式积累,并且分子量比游离 LC3B 高得多。当所有哺乳动物 LC3/GABARAP 同工型的预处理版本在 ATG4 缺陷细胞系中表达时,我们观察到类似的积累,这表明 LC3/GABARAP 也可以附着到其他较大的分子上。我们鉴定出 ATG3,即参与 LC3/GABARAP 脂质化的 E2 样酶,作为与多个 LC3/GABARAP 副本共轭的一个靶标。我们表明 LC3B-ATG3 缀合物与脂质化之前形成的 LC3B-ATG3 硫酯中间物不同,并且我们通过生化实验证明 ATG4B 可以切割 LC3B-ATG3 缀合物。最后,我们确定 ATG3 残基赖氨酸 243 为 LC3B 修饰位点。总的来说,我们提供了哺乳动物 LC3/GABARAP 在后翻译水平修饰蛋白质的第一个细胞证据,类似于泛素化(“LC3 化”),ATG4 蛋白酶作为去泛素化酶来抵消这种修饰(“去 LC3 化”)。