Krauss Veiko, Fassl Anne, Fiebig Petra, Patties Ina, Sass Heinz
Department of Genetics, Institute of Biology II, University of Leipzig, Johannisallee 21-23, 04103 Leipzig, Germany.
BMC Evol Biol. 2006 Mar 2;6:18. doi: 10.1186/1471-2148-6-18.
In eukaryotes, histone H3 lysine 9 (H3K9) methylation is a common mechanism involved in gene silencing and the establishment of heterochromatin. The loci of the major heterochromatic H3K9 methyltransferase Su(var)3-9 and the functionally unrelated gamma subunit of the translation initiation factor eIF2 are fused in Drosophila melanogaster. Here we examined the phylogenetic distribution of this unusual gene fusion and the molecular evolution of the H3K9 HMTase Su(var)3-9.
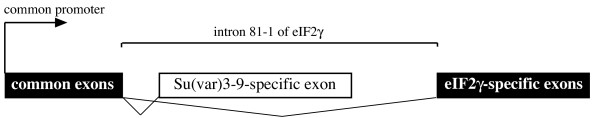
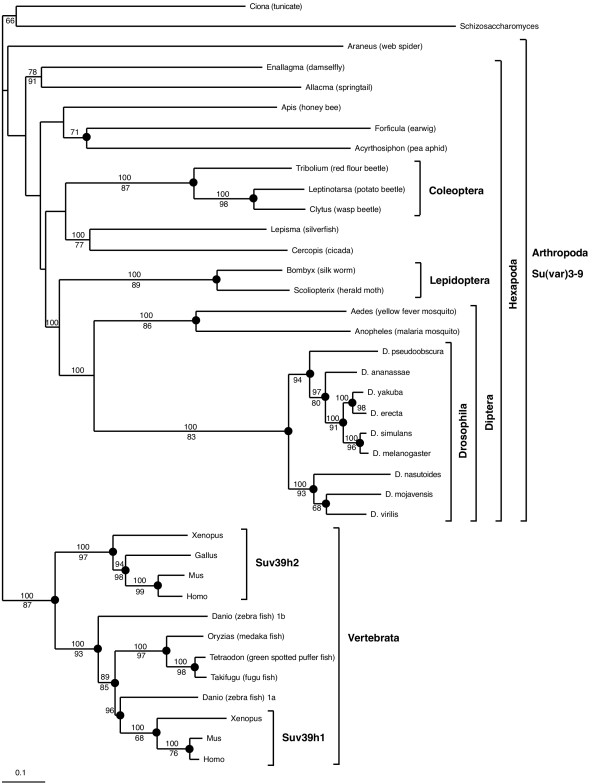
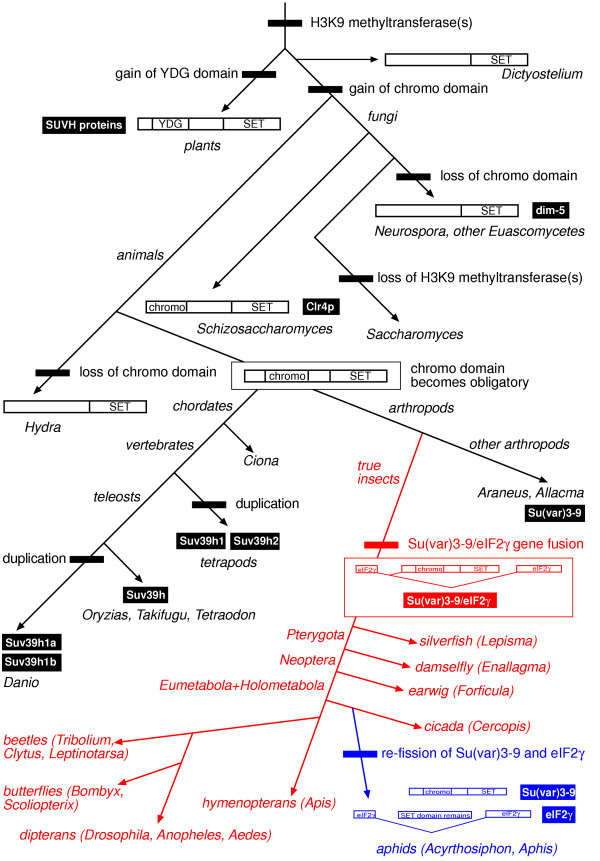
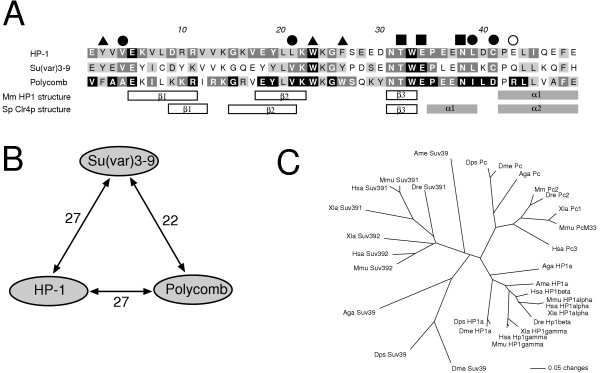
We show that the gene fusion had taken place in the ancestral line of winged insects and silverfishs (Dicondylia) about 400 million years ago. We cloned Su(var)3-9 genes from a collembolan and a spider where both genes ancestrally exist as independent transcription units. In contrast, we found a Su(var)3-9-specific exon inside the conserved intron position 81-1 of the eIF2gamma gene structure in species of eight different insect orders. Intriguinly, in the pea aphid Acyrthosiphon pisum, we detected only sequence remains of this Su(var)3-9 exon in the eIF2gamma intron, along with an eIF2gamma-independent Su(var)3-9 gene. This reveals an evolutionary re-fission of both genes in aphids. Su(var)3-9 chromo domains are similar to HP1 chromo domains, which points to a potential binding activity to methylated K9 of histone H3. SET domain comparisons suggest a weaker methyltransferase activity of Su(var)3-9 in comparison to other H3K9 HMTases. Astonishingly, 11 of 19 previously described, deleterious amino acid substitutions found in Drosophila Su(var)3-9 are seemingly compensable through accompanying substitutions during evolution.
Examination of the Su(var)3-9 evolution revealed strong evidence for the establishment of the Su(var)3-9/eIF2gamma gene fusion in an ancestor of dicondylic insects and a re-fission of this fusion during the evolution of aphids. Our comparison of 65 selected chromo domains and 93 selected SET domains from Su(var)3-9 and related proteins offers functional predictions concerning both domains in Su(var)3-9 proteins.
在真核生物中,组蛋白H3赖氨酸9(H3K9)甲基化是参与基因沉默和异染色质形成的常见机制。在果蝇中,主要的异染色质H3K9甲基转移酶Su(var)3-9的基因座与翻译起始因子eIF2功能无关的γ亚基融合。在此,我们研究了这种异常基因融合的系统发育分布以及H3K9组蛋白甲基转移酶Su(var)3-9的分子进化。
我们发现该基因融合发生在约4亿年前有翅昆虫和衣鱼(双髁类)的祖先谱系中。我们从弹尾虫和蜘蛛中克隆了Su(var)3-9基因,在这两种生物中,这两个基因在祖先中均作为独立的转录单元存在。相反,我们在八个不同昆虫目的物种的eIF2γ基因结构的保守内含子位置81-1内发现了一个Su(var)3-9特异性外显子。有趣的是,在豌豆蚜中,我们仅在eIF2γ内含子中检测到该Su(var)3-9外显子的序列残迹,以及一个独立于eIF2γ的Su(var)3-9基因。这揭示了蚜虫中这两个基因的进化性重新分裂。Su(var)3-9染色质结构域与HP1染色质结构域相似,这表明其对组蛋白H3甲基化的K9具有潜在的结合活性。SET结构域比较表明,与其他H3K9组蛋白甲基转移酶相比,Su(var)3-9的甲基转移酶活性较弱。令人惊讶的是,在果蝇Su(var)3-9中发现的19个先前描述的有害氨基酸取代中有11个似乎可通过进化过程中的伴随取代得到补偿。
对Su(var)3-9进化的研究揭示了强有力的证据,证明在双髁类昆虫的祖先中建立了Su(var)3-9/eIF2γ基因融合,并且在蚜虫的进化过程中该融合发生了重新分裂。我们对来自Su(var)3-9和相关蛋白的65个选定染色质结构域和93个选定SET结构域的比较提供了有关Su(var)3-9蛋白中这两个结构域的功能预测。