Loureiro Joana, Ploegh Hidde L
Whitehead Institute, 9 Cambridge Center, Cambridge, Massachusetts, USA.
Adv Immunol. 2006;92:225-305. doi: 10.1016/S0065-2776(06)92006-9.
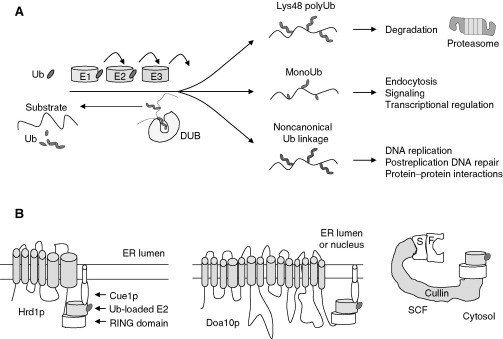
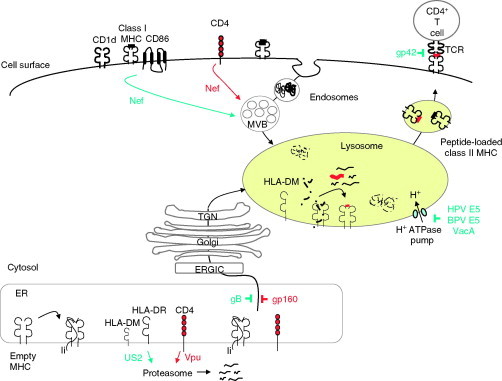
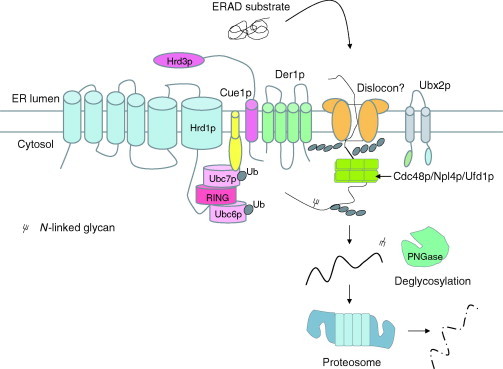
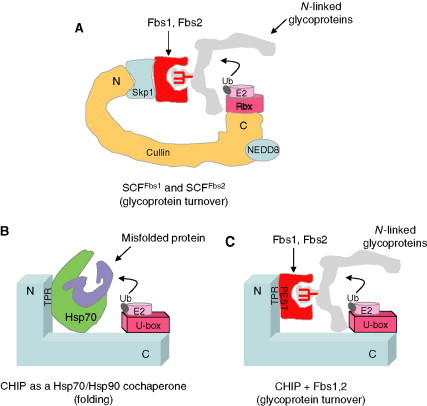
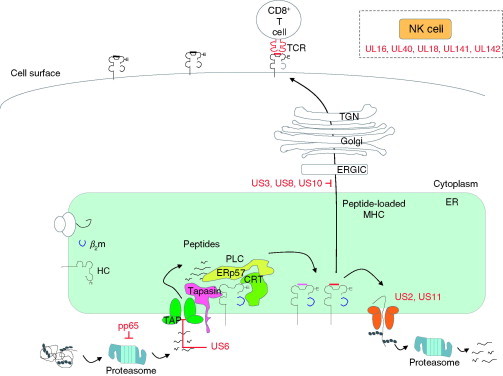
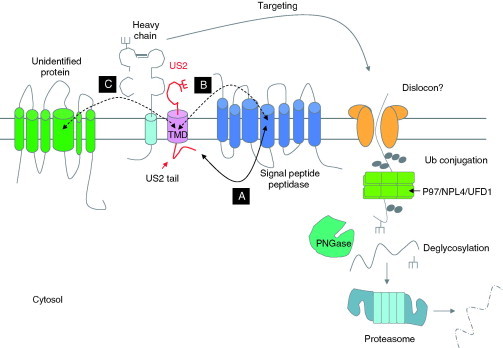
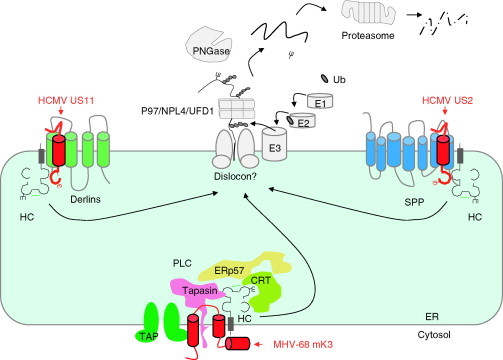
Relatively small genomes and high replication rates allow viruses and bacteria to accumulate mutations. This continuously presents the host immune system with new challenges. On the other side of the trenches, an increasingly well-adjusted host immune response, shaped by coevolutionary history, makes a pathogen's life a rather complicated endeavor. It is, therefore, no surprise that pathogens either escape detection or modulate the host immune response, often by redirecting normal cellular pathways to their advantage. For the purpose of this chapter, we focus mainly on the manipulation of the class I and class II major histocompatibility complex (MHC) antigen presentation pathways and the ubiquitin (Ub)-proteasome system by both viral and bacterial pathogens. First, we describe the general features of antigen presentation pathways and the Ub-proteasome system and then address how they are manipulated by pathogens. We discuss the many human cytomegalovirus (HCMV)-encoded immunomodulatory genes that interfere with antigen presentation (immunoevasins) and focus on the HCMV immunoevasins US2 and US11, which induce the degradation of class I MHC heavy chains by the proteasome by catalyzing their export from the endoplasmic reticulum (ER)-membrane into the cytosol, a process termed ER dislocation. US2- and US11-mediated subversion of ER dislocation ensures proteasomal degradation of class I MHC molecules and presumably allows HCMV to avoid recognition by cytotoxic T cells, whilst providing insight into general aspects of ER-associated degradation (ERAD) which is used by eukaryotic cells to purge their ER of defective proteins. We discuss the similarities and differences between the distinct pathways co-opted by US2 and US11 for dislocation and degradation of human class I MHC molecules and also a putatively distinct pathway utilized by the murine herpes virus (MHV)-68 mK3 immunoevasin for ER dislocation of murine class I MHC. We speculate on the implications of the three pathogen-exploited dislocation pathways to cellular ER quality control. Moreover, we discuss the ubiquitin (Ub)-proteasome system and its position at the core of antigen presentation as proteolysis and intracellular trafficking rely heavily on Ub-dependent processes. We add a few examples of manipulation of the Ub-proteasome system by pathogens in the context of the immune system and such diverse aspects of the host-pathogen relationship as virus budding, bacterial chromosome integration, and programmed cell death, to name a few. Finally, we speculate on newly found pathogen-encoded deubiquitinating enzymes (DUBs) and their putative roles in modulation of host-pathogen interactions.
相对较小的基因组和较高的复制率使病毒和细菌能够积累突变。这不断给宿主免疫系统带来新的挑战。在斗争的另一方,由共同进化历史塑造的、日益适应性良好的宿主免疫反应,使病原体的生存变得相当复杂。因此,病原体常常通过将正常细胞途径导向自身优势来逃避检测或调节宿主免疫反应,这并不奇怪。在本章中,我们主要关注病毒和细菌病原体对I类和II类主要组织相容性复合体(MHC)抗原呈递途径以及泛素(Ub)-蛋白酶体系统的操纵。首先,我们描述抗原呈递途径和Ub-蛋白酶体系统的一般特征,然后探讨它们是如何被病原体操纵的。我们讨论了许多人类巨细胞病毒(HCMV)编码的干扰抗原呈递的免疫调节基因(免疫逃避蛋白),并重点关注HCMV免疫逃避蛋白US2和US11,它们通过催化I类MHC重链从内质网(ER)膜转运到细胞质中,诱导蛋白酶体对其降解,这一过程称为ER错位。US2和US11介导的ER错位颠覆确保了I类MHC分子的蛋白酶体降解,可能使HCMV避免被细胞毒性T细胞识别,同时深入了解真核细胞用于清除内质网中缺陷蛋白的内质网相关降解(ERAD)的一般方面。我们讨论了US2和US11用于人类I类MHC分子错位和降解的不同途径之间的异同,以及小鼠疱疹病毒(MHV)-68 mK3免疫逃避蛋白用于小鼠I类MHC的ER错位的一种可能不同的途径。我们推测这三种病原体利用的错位途径对细胞内质网质量控制的影响。此外,我们讨论泛素(Ub)-蛋白酶体系统及其在抗原呈递核心位置的情况,因为蛋白水解和细胞内运输严重依赖于Ub依赖的过程。我们还列举了病原体在免疫系统背景下操纵Ub-蛋白酶体系统的一些例子,以及宿主-病原体关系的其他不同方面,如病毒出芽、细菌染色体整合和程序性细胞死亡等。最后,我们推测新发现的病原体编码的去泛素化酶(DUBs)及其在调节宿主-病原体相互作用中的假定作用。