Jékely Gáspár
European Molecular Biology Laboratory, Meyerhofstrasse 1, 69117 Heidelberg, Germany.
Biol Direct. 2007 Jan 19;2:3. doi: 10.1186/1745-6150-2-3.
The origin of eukaryotic cells was one of the most dramatic evolutionary transitions in the history of life. It is generally assumed that eukaryotes evolved later then prokaryotes by the transformation or fusion of prokaryotic lineages. However, as yet there is no consensus regarding the nature of the prokaryotic group(s) ancestral to eukaryotes. Regardless of this, a hardly debatable fundamental novel characteristic of the last eukaryotic common ancestor was the ability to exploit prokaryotic biomass by the ingestion of entire cells, i.e. phagocytosis. The recent advances in our understanding of the social life of prokaryotes may help to explain the origin of this form of total exploitation.
Here I propose that eukaryotic cells originated in a social environment, a differentiated microbial mat or biofilm that was maintained by the cooperative action of its members. Cooperation was costly (e.g. the production of developmental signals or an extracellular matrix) but yielded benefits that increased the overall fitness of the social group. I propose that eukaryotes originated as selfish cheaters that enjoyed the benefits of social aggregation but did not contribute to it themselves. The cheaters later evolved into predators that lysed other cells and eventually became professional phagotrophs. During several cycles of social aggregation and dispersal the number of cheaters was contained by a chicken game situation, i.e. reproductive success of cheaters was high when they were in low abundance but was reduced when they were over-represented. Radical changes in cell structure, including the loss of the rigid prokaryotic cell wall and the development of endomembranes, allowed the protoeukaryotes to avoid cheater control and to exploit nutrients more efficiently. Cellular changes were buffered by both the social benefits and the protective physico-chemical milieu of the interior of biofilms. Symbiosis with the mitochondial ancestor evolved after phagotrophy as alphaproteobacterial prey developed post-ingestion defence mechanisms to circumvent digestion in the food vacuole. Mitochondrial symbiosis triggered the origin of the nucleus. Cilia evolved last and allowed eukaryotes to predate also on planktonic prey. I will discuss how this scenario may possibly fit into the contrasting phylogenetic frameworks that have been proposed.
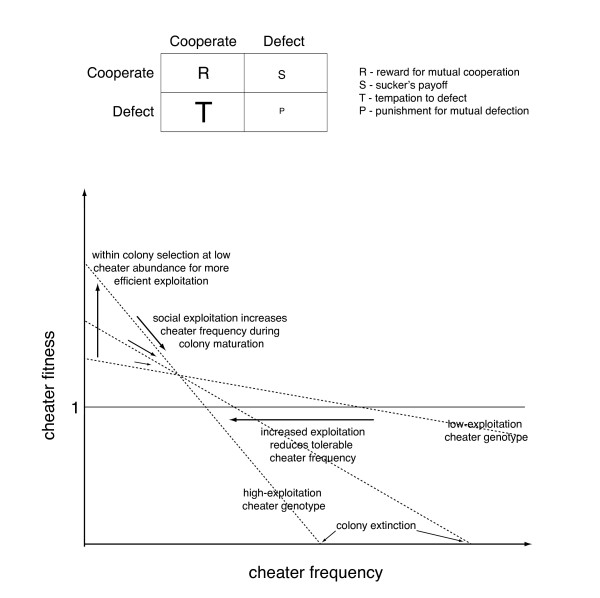
Some aspects of the hypothesis can be tested experimentally by studying the level of exploitation cheaters can reach in social microbes. It would be interesting to test whether absorption of nutrients from lysed fellow colony members can happen and if cheaters can evolve into predators that actively digest neighbouring cells.
The hypothesis highlights the importance of social exploitation in cell evolution and how a social environment can buffer drastic cellular transformations that would be lethal for planktonic forms.
真核细胞的起源是生命史上最重大的进化转变之一。一般认为,真核生物是在原核生物之后通过原核生物谱系的转化或融合而进化的。然而,关于真核生物祖先的原核生物群体的性质,目前尚无共识。尽管如此,最后一个真核生物共同祖先几乎无可争议的一个基本新特征是通过摄取整个细胞来利用原核生物生物质的能力,即吞噬作用。我们对原核生物社会生活理解的最新进展可能有助于解释这种全面利用形式的起源。
在此我提出,真核细胞起源于一个社会环境,即一个由其成员的协同作用维持的分化微生物垫或生物膜。合作成本高昂(例如发育信号或细胞外基质的产生),但带来的益处增加了社会群体的整体适应性。我提出真核生物起源于自私的作弊者,它们享受社会聚集的益处但自身并不为此做出贡献。这些作弊者后来进化成捕食者,裂解其他细胞,最终成为专业吞噬者。在社会聚集和分散的几个循环中,作弊者的数量受到一种斗鸡博弈情况的限制,即作弊者数量较少时繁殖成功率高,但数量过多时则会降低。细胞结构的剧烈变化,包括刚性原核细胞壁的丧失和内膜系统的发展,使原始真核生物能够避免被作弊者控制,并更有效地利用营养物质。细胞变化受到生物膜内部的社会效益和保护性物理化学环境的缓冲。吞噬作用之后,随着α-变形菌猎物在被摄入后发展出防御机制以规避食物泡中的消化,与线粒体祖先的共生关系得以进化。线粒体共生引发了细胞核的起源。纤毛是最后进化出来的,使真核生物也能够捕食浮游猎物。我将讨论这个假说如何可能符合已提出的不同系统发育框架。
该假说的某些方面可以通过研究作弊者在社会微生物中能够达到的利用水平进行实验检验。测试从裂解的同菌落成员中吸收营养物质是否会发生以及作弊者是否能进化成主动消化邻近细胞的捕食者将会很有趣。
该假说强调了社会利用在细胞进化中的重要性,以及社会环境如何能够缓冲对浮游形式致命的剧烈细胞转变。