Evolutionary Systems Research Group, Institute of Evolution, Centre for Ecological Research, Klebelsberg Kunó str. 3., Tihany, 8237, Hungary.
MTA-ELTE Theoretical Biology and Evolutionary Ecology Research Group, Department of Plant Taxonomy and Ecology, Eötvös Loránd University, Pázmány Péter sétány 1/c, Budapest, 1117, Hungary.
Cell Mol Life Sci. 2020 Sep;77(18):3503-3523. doi: 10.1007/s00018-020-03462-6. Epub 2020 Feb 1.
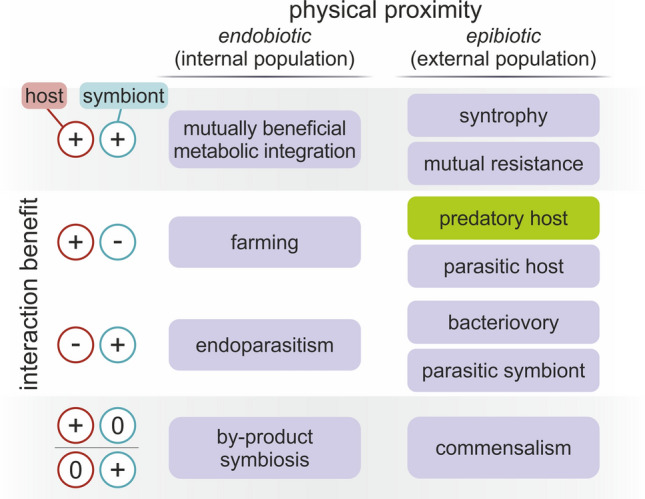
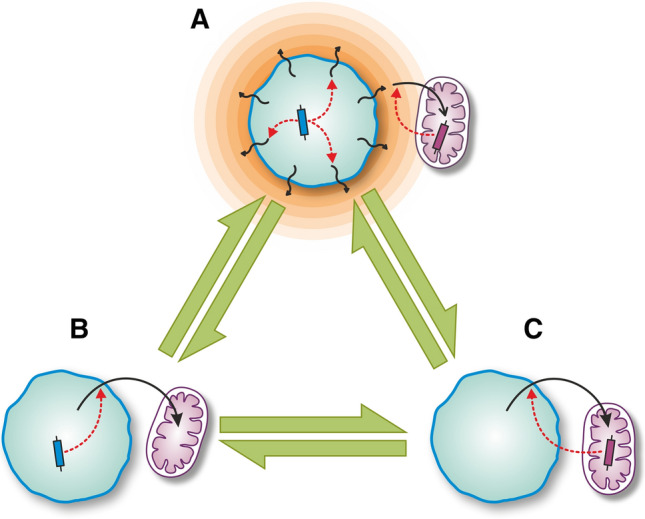
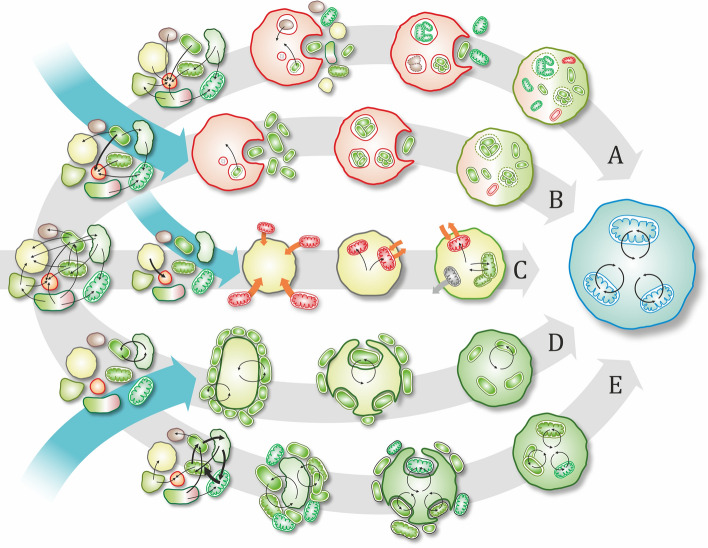
Endosymbiosis and organellogenesis are virtually unknown among prokaryotes. The single presumed example is the endosymbiogenetic origin of mitochondria, which is hidden behind the event horizon of the last eukaryotic common ancestor. While eukaryotes are monophyletic, it is unlikely that during billions of years, there were no other prokaryote-prokaryote endosymbioses as symbiosis is extremely common among prokaryotes, e.g., in biofilms. Therefore, it is even more precarious to draw conclusions about potentially existing (or once existing) prokaryotic endosymbioses based on a single example. It is yet unknown if the bacterial endosymbiont was captured by a prokaryote or by a (proto-)eukaryote, and if the process of internalization was parasitic infection, slow engulfment, or phagocytosis. In this review, we accordingly explore multiple mechanisms and processes that could drive the evolution of unicellular microbial symbioses with a special attention to prokaryote-prokaryote interactions and to the mitochondrion, possibly the single prokaryotic endosymbiosis that turned out to be a major evolutionary transition. We investigate the ecology and evolutionary stability of inter-species microbial interactions based on dependence, physical proximity, cost-benefit budget, and the types of benefits, investments, and controls. We identify challenges that had to be conquered for the mitochondrial host to establish a stable eukaryotic lineage. Any assumption about the initial interaction of the mitochondrial ancestor and its contemporary host based solely on their modern relationship is rather perilous. As a result, we warn against assuming an initial mutually beneficial interaction based on modern mitochondria-host cooperation. This assumption is twice fallacious: (i) endosymbioses are known to evolve from exploitative interactions and (ii) cooperativity does not necessarily lead to stable mutualism. We point out that the lack of evidence so far on the evolution of endosymbiosis from mutual syntrophy supports the idea that mitochondria emerged from an exploitative (parasitic or phagotrophic) interaction rather than from syntrophy.
内共生和细胞器发生在原核生物中几乎是未知的。唯一被假定的例子是线粒体的内共生起源,它隐藏在最后一个真核生物共同祖先的事件视界之后。虽然真核生物是单系的,但在数十亿年的时间里,不可能没有其他原核生物-原核生物的内共生,因为共生在原核生物中非常普遍,例如在生物膜中。因此,基于单一例子来推断潜在存在(或曾经存在)的原核内共生关系更加危险。目前还不清楚细菌内共生体是被原核生物还是(原)真核生物捕获的,如果内化过程是寄生感染、缓慢吞噬还是吞噬作用。在这篇综述中,我们相应地探索了多种可能导致单细胞微生物共生进化的机制和过程,特别关注原核生物-原核生物的相互作用和线粒体,线粒体可能是唯一被证明是重大进化转折点的原核内共生体。我们根据依赖性、物理接近度、成本效益预算以及收益、投资和控制的类型,研究了种间微生物相互作用的生态学和进化稳定性。我们确定了线粒体宿主为建立稳定的真核谱系而必须克服的挑战。任何仅基于现代关系对线粒体祖先及其当代宿主最初相互作用的假设都是相当危险的。因此,我们警告不要仅基于现代线粒体-宿主合作就假设最初的互利相互作用。这种假设是双重错误的:(i)内共生是从剥削性相互作用中进化而来的,(ii)合作不一定会导致稳定的共生关系。我们指出,目前关于内共生从互利共栖进化而来的证据不足,这支持了线粒体从剥削性(寄生或吞噬)相互作用而不是从共栖中出现的观点。