Department of Physiology and Biophysics, University of Washington, Seattle, WA 98195, USA.
J Gen Physiol. 2010 Feb;135(2):99-114. doi: 10.1085/jgp.200910345.
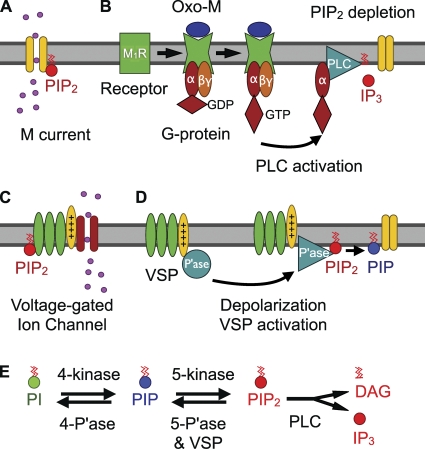
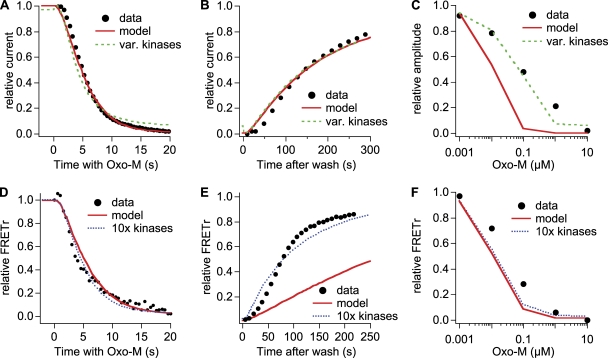
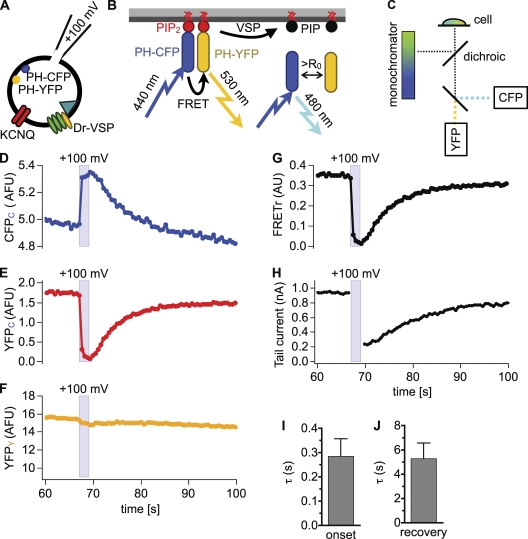
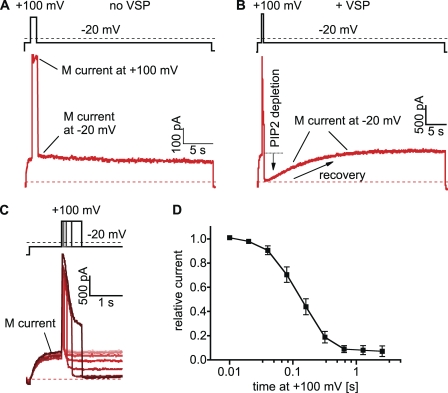
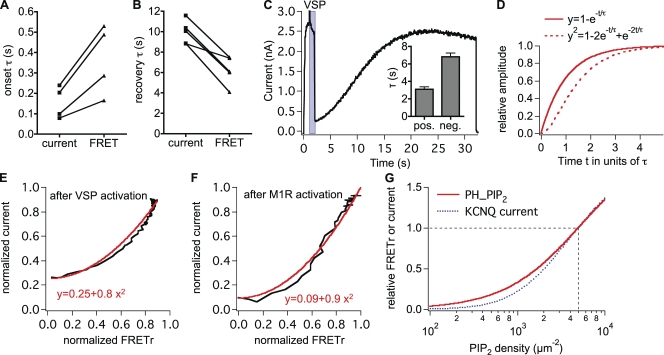
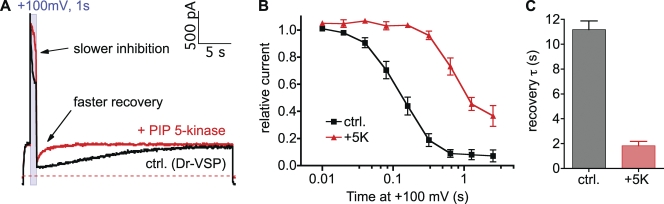
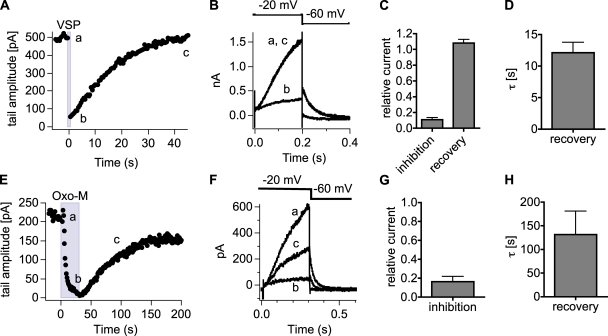
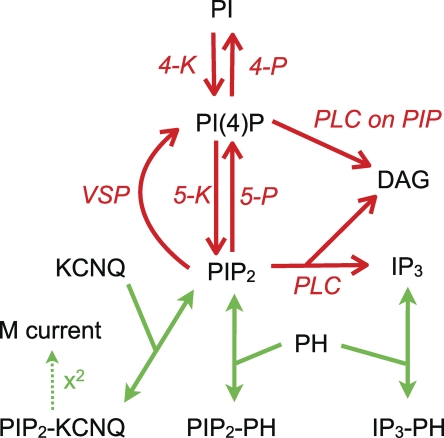
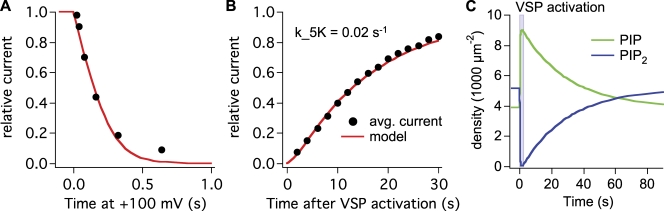
The signaling phosphoinositide phosphatidylinositol 4,5-bisphosphate (PIP(2)) is synthesized in two steps from phosphatidylinositol by lipid kinases. It then interacts with KCNQ channels and with pleckstrin homology (PH) domains among many other physiological protein targets. We measured and developed a quantitative description of these metabolic and protein interaction steps by perturbing the PIP(2) pool with a voltage-sensitive phosphatase (VSP). VSP can remove the 5-phosphate of PIP(2) with a time constant of tau <300 ms and fully inhibits KCNQ currents in a similar time. PIP(2) was then resynthesized from phosphatidylinositol 4-phosphate (PIP) quickly, tau = 11 s. In contrast, resynthesis of PIP(2) after activation of phospholipase C by muscarinic receptors took approximately 130 s. These kinetic experiments showed that (1) PIP(2) activation of KCNQ channels obeys a cooperative square law, (2) the PIP(2) residence time on channels is <10 ms and the exchange time on PH domains is similarly fast, and (3) the step synthesizing PIP(2) by PIP 5-kinase is fast and limited primarily by a step(s) that replenishes the pool of plasma membrane PI(4)P. We extend the kinetic model for signaling from M(1) muscarinic receptors, presented in our companion paper in this issue (Falkenburger et al. 2010. J. Gen. Physiol. doi:10.1085/jgp.200910344), with this new information on PIP(2) synthesis and KCNQ interaction.
信号磷酸肌醇 4,5-二磷酸(PIP(2))由脂质激酶从肌醇磷脂两步合成。然后,它与 KCNQ 通道相互作用,并与许多其他生理蛋白靶标中的pleckstrin 同源(PH)结构域相互作用。我们通过电压敏感磷酸酶(VSP)对 PIP(2)池进行扰动,测量并对这些代谢和蛋白相互作用步骤进行了定量描述。VSP 可以在 <300 ms 的时间常数内去除 PIP(2)的 5-磷酸,并且在相似的时间内完全抑制 KCNQ 电流。然后,PIP(2)从肌醇 4-磷酸(PIP)快速重新合成,tau = 11 s。相比之下,通过毒蕈碱受体激活磷脂酶 C 后,PIP(2)的重新合成大约需要 130 s。这些动力学实验表明:(1) KCNQ 通道的 PIP(2)激活符合协同平方律;(2) PIP(2)在通道上的停留时间<10 ms,在 PH 结构域上的交换时间也同样快;(3) PIP 5-激酶合成 PIP(2)的步骤很快,主要受补充质膜 PI(4)P 池的步骤限制。我们将本期(Falkenburger 等人,2010 年。J. Gen. Physiol.,doi:10.1085/jgp.200910344)中报道的 M(1)毒蕈碱受体信号转导的动力学模型扩展为包括我们的新信息PIP(2)合成和 KCNQ 相互作用。