Instituto de Biomedicina de Valencia, Consejo Superior de Investigaciones Científicas (IBV-CSIC), Valencia, Spain.
BMC Evol Biol. 2010 Feb 22;10:56. doi: 10.1186/1471-2148-10-56.
HECT ubiquitin ligases (HECT E3s) are key components of the eukaryotic ubiquitin-proteasome system and are involved in the genesis of several human diseases. In this study, I analyze the patterns of diversification of HECT E3s since animals emerged in order to provide the right framework to understand the functional data available for proteins of this family.
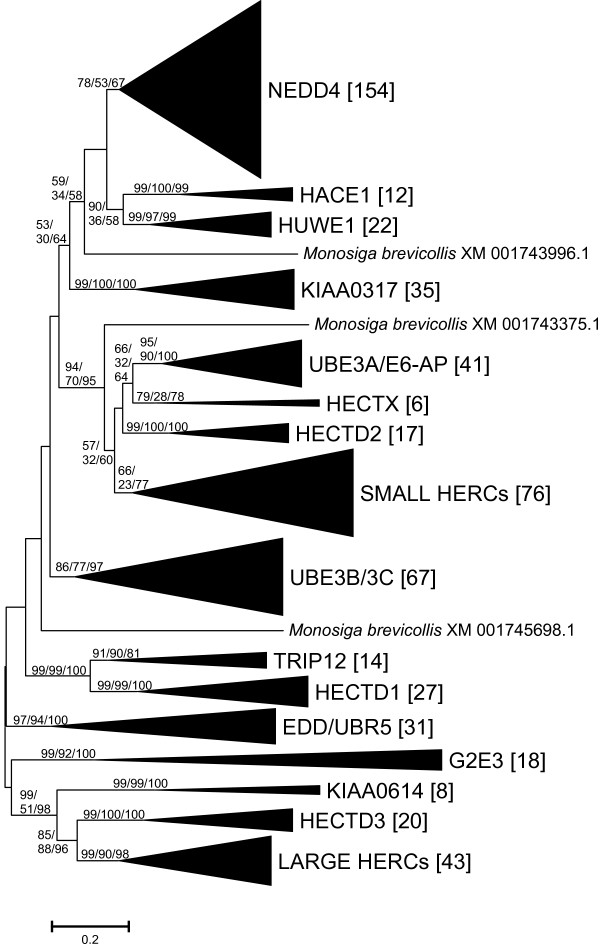
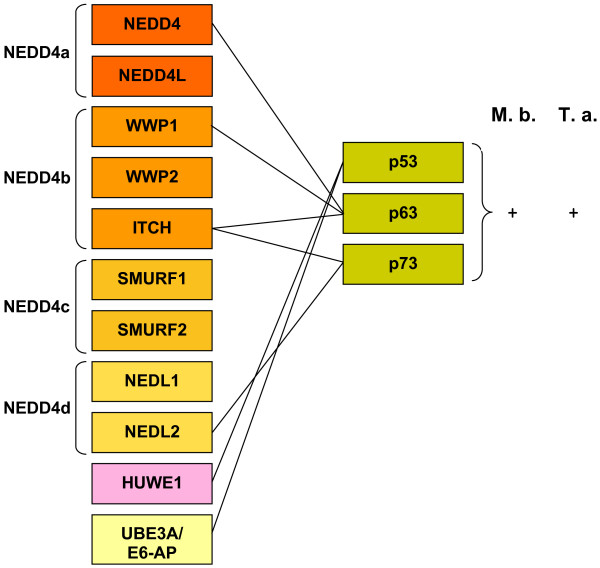
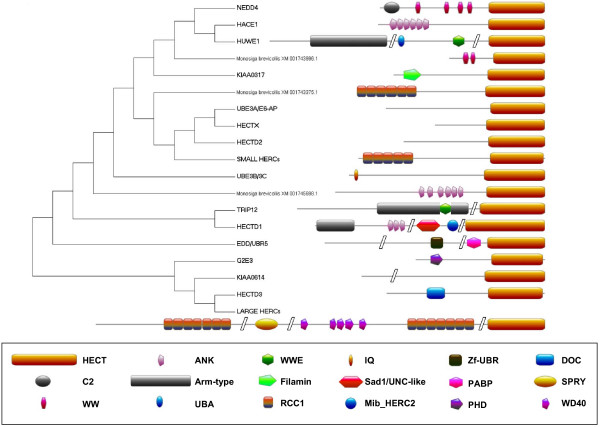
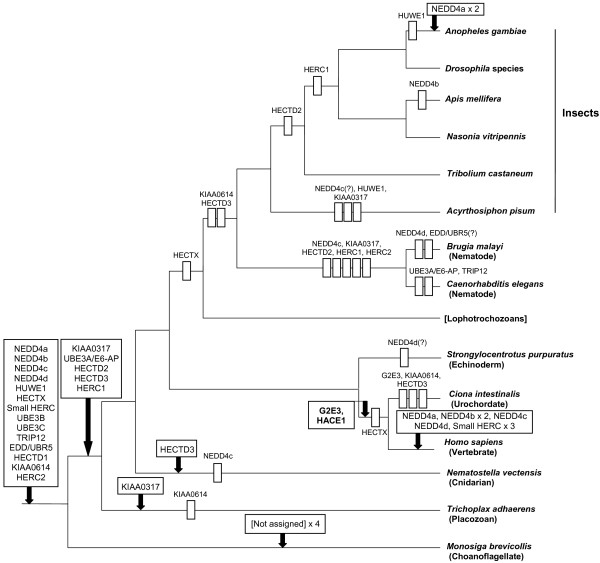
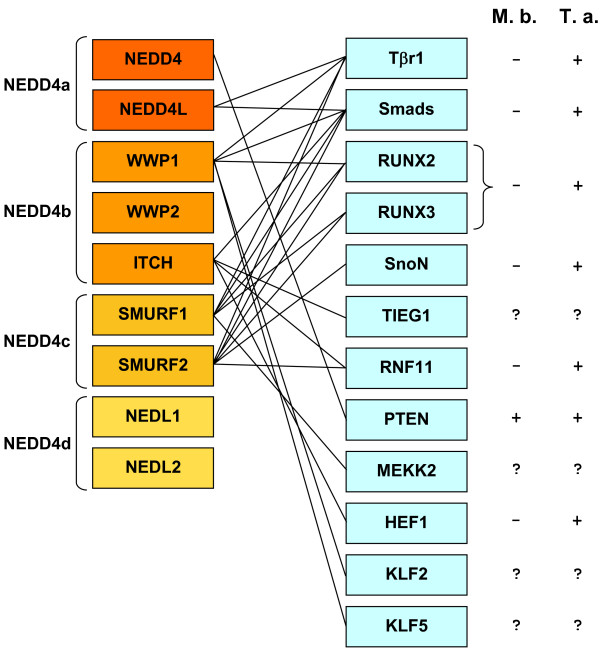
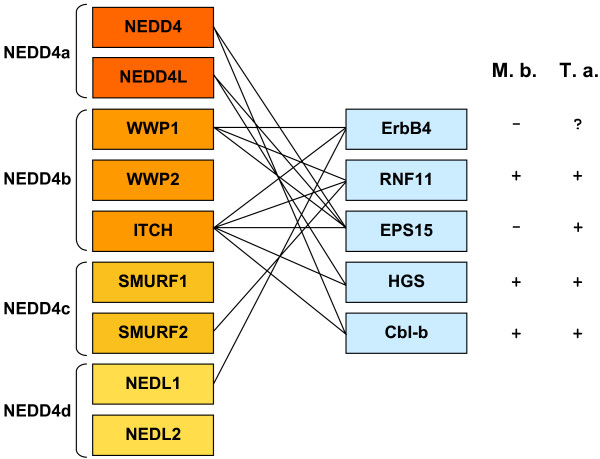
I show that the current classification of HECT E3s into three groups (NEDD4-like E3s, HERCs and single-HECT E3s) is fundamentally incorrect. First, the existence of a "Single-HECT E3s" group is not supported by phylogenetic analyses. Second, the HERC proteins must be divided into two subfamilies (Large HERCs, Small HERCs) that are evolutionarily very distant, their structural similarity being due to convergence and not to a common origin. Sequence and structural analyses show that animal HECT E3s can be naturally classified into 16 subfamilies. Almost all of them appeared either before animals originated or in early animal evolution. More recently, multiple gene losses have occurred independently in some lineages (nematodes, insects, urochordates), the same groups that have also lost genes of another type of E3s (RBR family). Interestingly, the emergence of some animal HECT E3s precedes the origin of key cellular systems that they regulate (TGF-beta and EGF signal transduction pathways; p53 family of transcription factors) and it can be deduced that distantly related HECT proteins have been independently co-opted to perform similar roles. This may contribute to explain why distantly related HECT E3s are involved in the genesis of multiple types of cancer.
The complex evolutionary history of HECT ubiquitin ligases in animals has been deciphered. The most appropriate model animals to study them and new theoretical and experimental lines of research are suggested by these results.
HECT 泛素连接酶(HECT E3s)是真核生物泛素蛋白酶体系统的关键组成部分,参与了几种人类疾病的发生。在本研究中,我分析了动物出现以来 HECT E3s 的多样化模式,以便为理解该家族蛋白的现有功能数据提供正确的框架。
我表明,目前将 HECT E3s 分为三组(NEDD4 样 E3s、HERCs 和单 HECT E3s)的分类方法是完全不正确的。首先,系统发育分析不支持“单 HECT E3s”组的存在。其次,HERC 蛋白必须分为两个亚家族(大 HERCs、小 HERCs),它们在进化上非常遥远,它们的结构相似是由于趋同而不是共同起源。序列和结构分析表明,动物 HECT E3s 可以自然地分为 16 个亚家族。几乎所有的亚家族都出现在动物起源之前或动物早期进化过程中。最近,一些谱系(线虫、昆虫、尾索动物)独立地发生了多次基因丢失,而这些谱系也失去了另一种 E3s(RBR 家族)的基因。有趣的是,一些动物 HECT E3s 的出现早于它们调节的关键细胞系统(TGF-β和 EGF 信号转导途径;p53 家族转录因子)的起源,可以推断出远缘的 HECT 蛋白已被独立地重新用于执行类似的功能。这可能有助于解释为什么远缘的 HECT E3s 参与了多种类型的癌症的发生。
动物中 HECT 泛素连接酶的复杂进化历史已经被揭示。这些结果为研究它们提供了最合适的模式动物和新的理论和实验研究方向。