Department of Microbiology and Immunology, University of Arkansas for Medical Sciences, Little Rock, Arkansas, USA.
PLoS One. 2010 May 24;5(5):e10790. doi: 10.1371/journal.pone.0010790.
The accessory gene regulator (agr) and staphylococcal accessory regulator (sarA) play opposing roles in Staphylococcus aureus biofilm formation. There is mounting evidence to suggest that these opposing roles are therapeutically relevant in that mutation of agr results in increased biofilm formation and decreased antibiotic susceptibility while mutation of sarA has the opposite effect. To the extent that induction of agr or inhibition of sarA could potentially be used to limit biofilm formation, this makes it important to understand the epistatic relationships between these two loci.
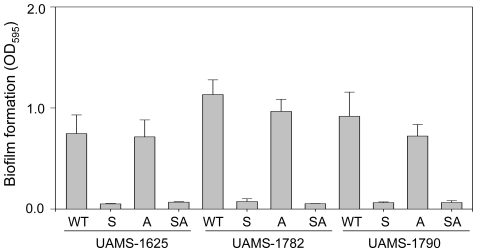
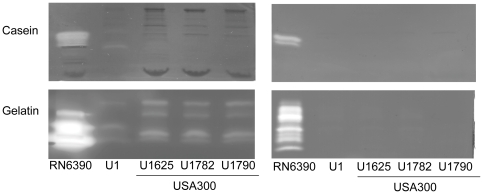
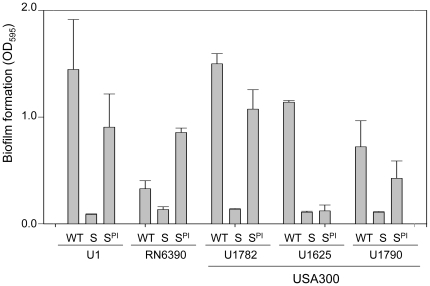
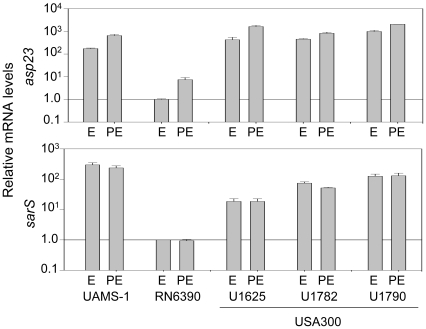
METHODOLOGY/PRINCIPAL FINDINGS: We generated isogenic sarA and agr mutants in clinical isolates of S. aureus and assessed the relative impact on biofilm formation. Mutation of agr resulted in an increased capacity to form a biofilm in the 8325-4 laboratory strain RN6390 but had little impact in clinical isolates S. aureus. In contrast, mutation of sarA resulted in a reduced capacity to form a biofilm in all clinical isolates irrespective of the functional status of agr. This suggests that the regulatory role of sarA in biofilm formation is independent of the interaction between sarA and agr and that sarA is epistatic to agr in this context. This was confirmed by demonstrating that restoration of sarA function restored the ability to form a biofilm even in the corresponding agr mutants. Mutation of sarA in clinical isolates also resulted in increased production of extracellular proteases and extracellular nucleases, both of which contributed to the biofilm-deficient phenotype of sarA mutants. However, studies comparing different strains with and without proteases inhibitors and/or mutation of the nuclease genes demonstrated that the agr-independent, sarA-mediated repression of extracellular proteases plays a primary role in this regard.
The results we report suggest that inhibitors of sarA-mediated regulation could be used to limit biofilm formation in S. aureus and that the efficacy of such inhibitors would not be limited by spontaneous mutation of agr in the human host.
辅助基因调控器(agr)和葡萄球菌辅助调节因子(sarA)在金黄色葡萄球菌生物膜形成中起着相反的作用。越来越多的证据表明,这些相反的作用在治疗上是相关的,因为 agr 的突变导致生物膜形成增加和抗生素敏感性降低,而 sarA 的突变则产生相反的效果。在诱导 agr 或抑制 sarA 可能被用来限制生物膜形成的程度上,这使得理解这两个基因座之间的上位关系变得非常重要。
方法/主要发现:我们在金黄色葡萄球菌的临床分离株中生成了同源的 sarA 和 agr 突变体,并评估了它们对生物膜形成的相对影响。agr 的突变导致 8325-4 实验室菌株 RN6390 的生物膜形成能力增加,但对临床分离株金黄色葡萄球菌影响不大。相比之下,sarA 的突变导致所有临床分离株的生物膜形成能力降低,而与 agr 的功能状态无关。这表明 sarA 在生物膜形成中的调节作用独立于 sarA 和 agr 之间的相互作用,并且在这种情况下 sarA 对 agr 具有上位性。通过证明恢复 sarA 的功能甚至可以在相应的 agr 突变体中恢复生物膜形成能力,证实了这一点。sarA 在临床分离株中的突变也导致细胞外蛋白酶和细胞外核酸酶的产量增加,这两者都导致了 sarA 突变体生物膜缺陷表型。然而,比较具有和不具有蛋白酶抑制剂的不同菌株以及 nuclease 基因的突变的研究表明,agr 独立的、sarA 介导的细胞外蛋白酶的抑制在这方面起着主要作用。
我们报告的结果表明,sarA 介导的调节抑制剂可以用于限制金黄色葡萄球菌的生物膜形成,并且此类抑制剂的功效不会受到宿主中人 agr 自发突变的限制。