Institute of Molecular Biophysics, Florida State University, Tallahassee, Florida, United States of America.
PLoS One. 2010 Sep 9;5(9):e12643. doi: 10.1371/journal.pone.0012643.
Isometric muscle contraction, where force is generated without muscle shortening, is a molecular traffic jam in which the number of actin-attached motors is maximized and all states of motor action are trapped with consequently high heterogeneity. This heterogeneity is a major limitation to deciphering myosin conformational changes in situ.
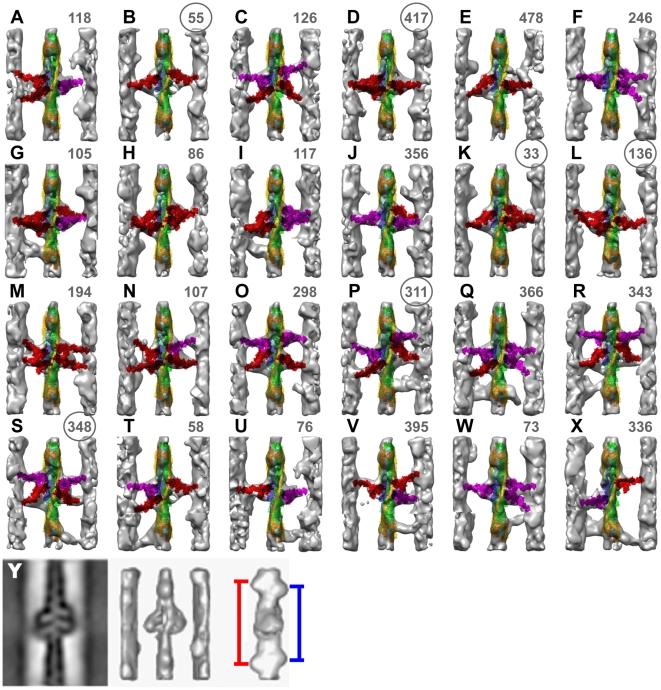
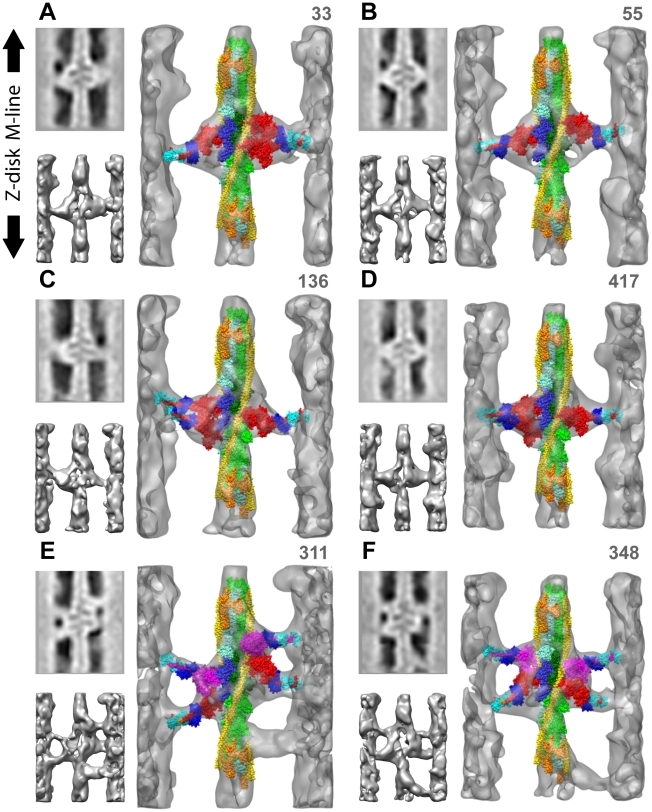
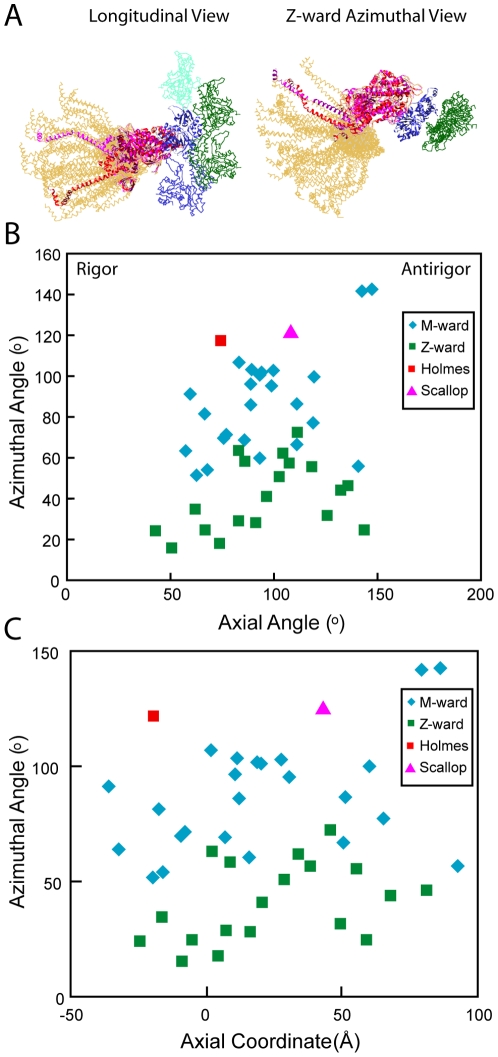
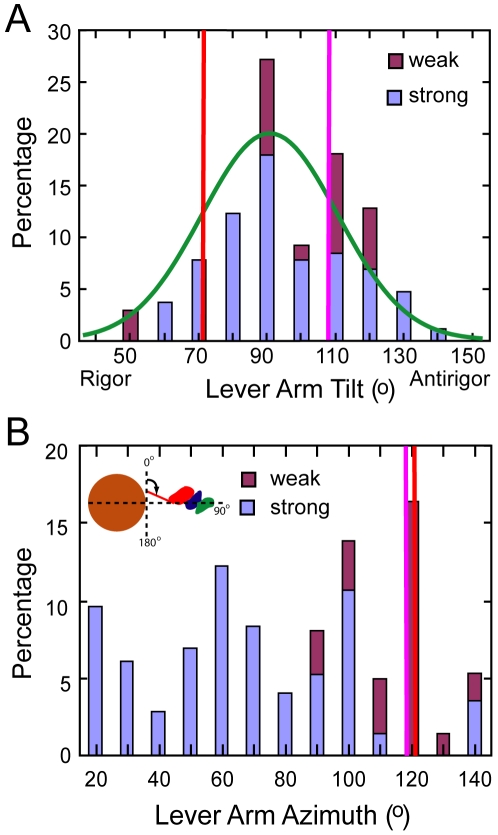
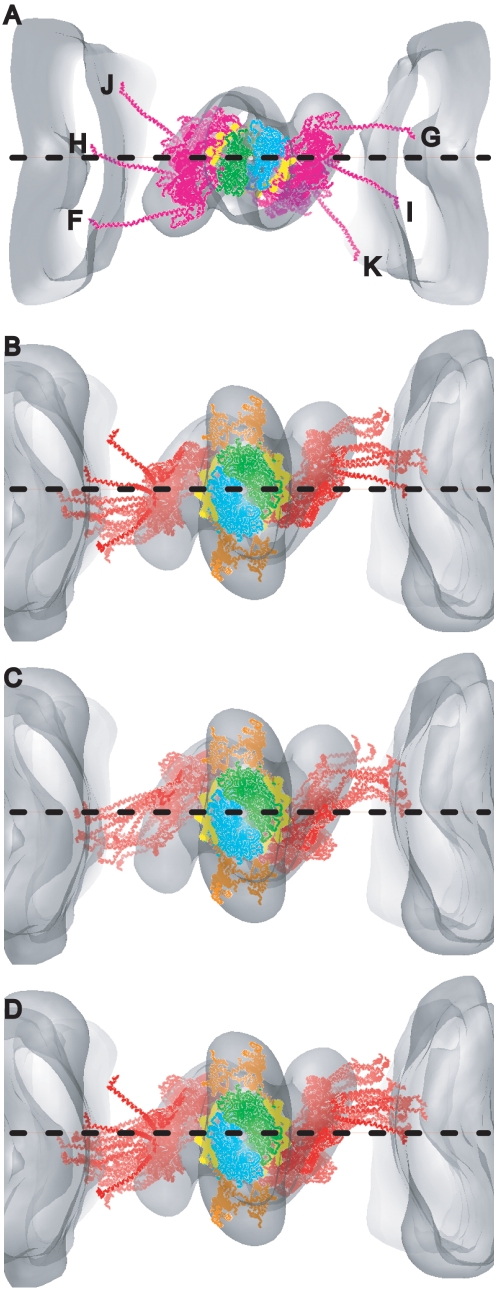
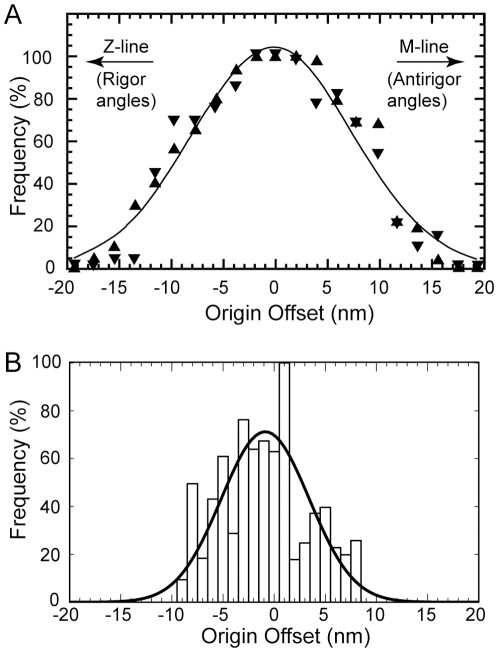
We used multivariate data analysis to group repeat segments in electron tomograms of isometrically contracting insect flight muscle, mechanically monitored, rapidly frozen, freeze substituted, and thin sectioned. Improved resolution reveals the helical arrangement of F-actin subunits in the thin filament enabling an atomic model to be built into the thin filament density independent of the myosin. Actin-myosin attachments can now be assigned as weak or strong by their motor domain orientation relative to actin. Myosin attachments were quantified everywhere along the thin filament including troponin. Strong binding myosin attachments are found on only four F-actin subunits, the "target zone", situated exactly midway between successive troponin complexes. They show an axial lever arm range of 77°/12.9 nm. The lever arm azimuthal range of strong binding attachments has a highly skewed, 127° range compared with X-ray crystallographic structures. Two types of weak actin attachments are described. One type, found exclusively in the target zone, appears to represent pre-working-stroke intermediates. The other, which contacts tropomyosin rather than actin, is positioned M-ward of the target zone, i.e. the position toward which thin filaments slide during shortening.
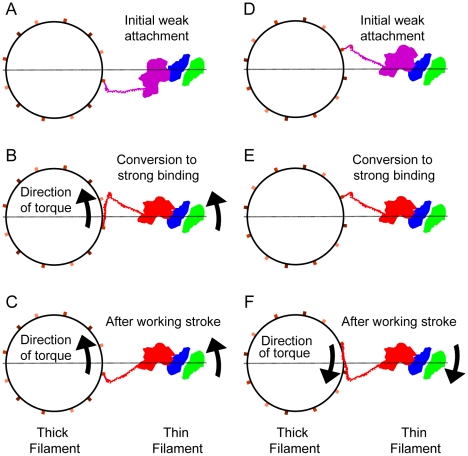
We present a model for the weak to strong transition in the myosin ATPase cycle that incorporates azimuthal movements of the motor domain on actin. Stress/strain in the S2 domain may explain azimuthal lever arm changes in the strong binding attachments. The results support previous conclusions that the weak attachments preceding force generation are very different from strong binding attachments.
等长肌肉收缩是一种分子交通堵塞,其中附着在肌动蛋白上的肌球蛋白数量达到最大值,并且所有肌球蛋白的运动状态都被捕获,从而导致高度异质性。这种异质性是解析肌球蛋白构象变化的主要限制。
我们使用多元数据分析将重复片段分组,这些片段来自等长收缩昆虫飞行肌的电子断层图像,该肌由机械监测,快速冷冻,冷冻替代和超薄切片。分辨率的提高揭示了 F-肌动蛋白亚基在薄丝中的螺旋排列,使原子模型能够独立于肌球蛋白构建到薄丝密度中。现在可以根据肌球蛋白相对于肌动蛋白的马达域取向将肌动球蛋白附着分配为弱或强。在薄丝的任何地方都可以定量肌球蛋白附着,包括肌钙蛋白。仅在四个 F-肌动蛋白亚基上发现强结合的肌球蛋白附着,称为“靶区”,正好位于连续肌钙蛋白复合物的中间。它们显示出 77°/12.9nm 的轴向杠杆臂范围。强结合附着的杠杆臂方位范围与 X 射线晶体结构相比具有高度倾斜的 127°范围。描述了两种类型的弱肌动球蛋白附着。一种类型仅在靶区中发现,似乎代表预工作冲程中间体。另一种类型与肌球蛋白接触而不是肌动蛋白,位于靶区的 M 向,即在缩短过程中薄丝滑动的位置。
我们提出了一个肌球蛋白 ATP 酶循环中从弱结合到强结合的转变模型,该模型结合了肌球蛋白在肌动蛋白上的方位运动。S2 结构域中的应力/应变可能解释了强结合附着的方位杠杆臂变化。结果支持先前的结论,即在产生力之前的弱附着与强结合附着非常不同。