Department of Chemistry, Johns Hopkins University, Baltimore, MD 21218, USA.
J Mol Biol. 2010 Nov 19;404(1):138-57. doi: 10.1016/j.jmb.2010.09.043. Epub 2010 Sep 25.
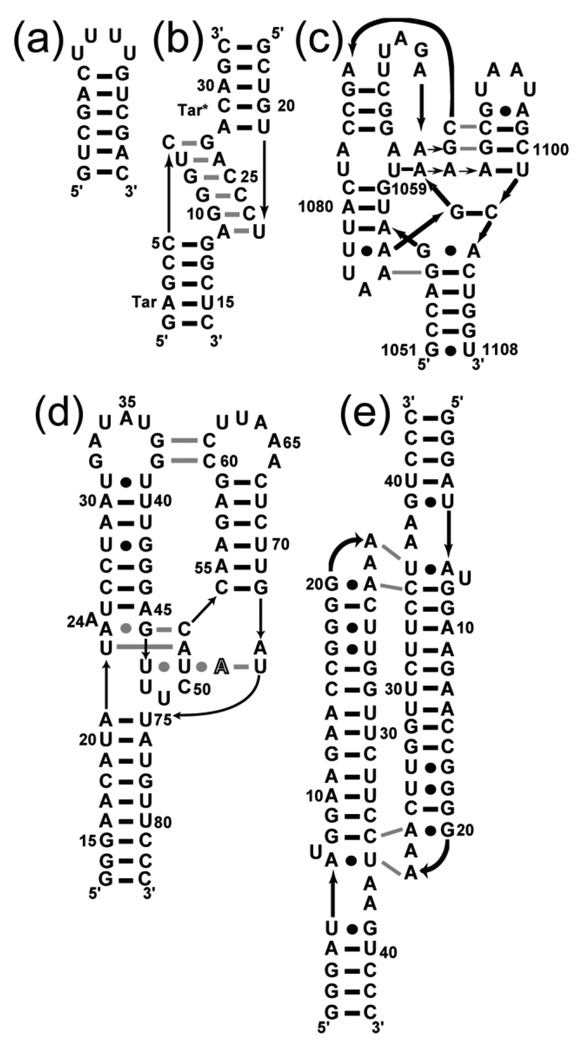
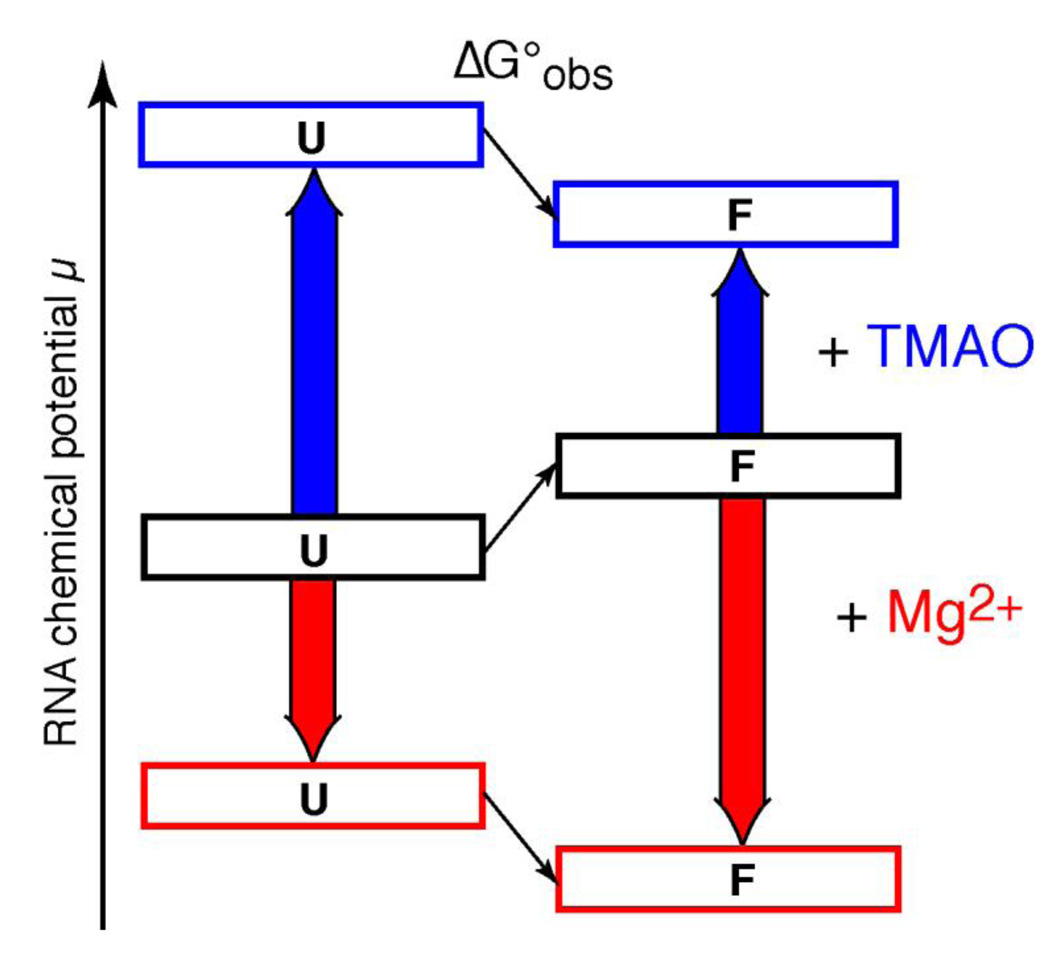
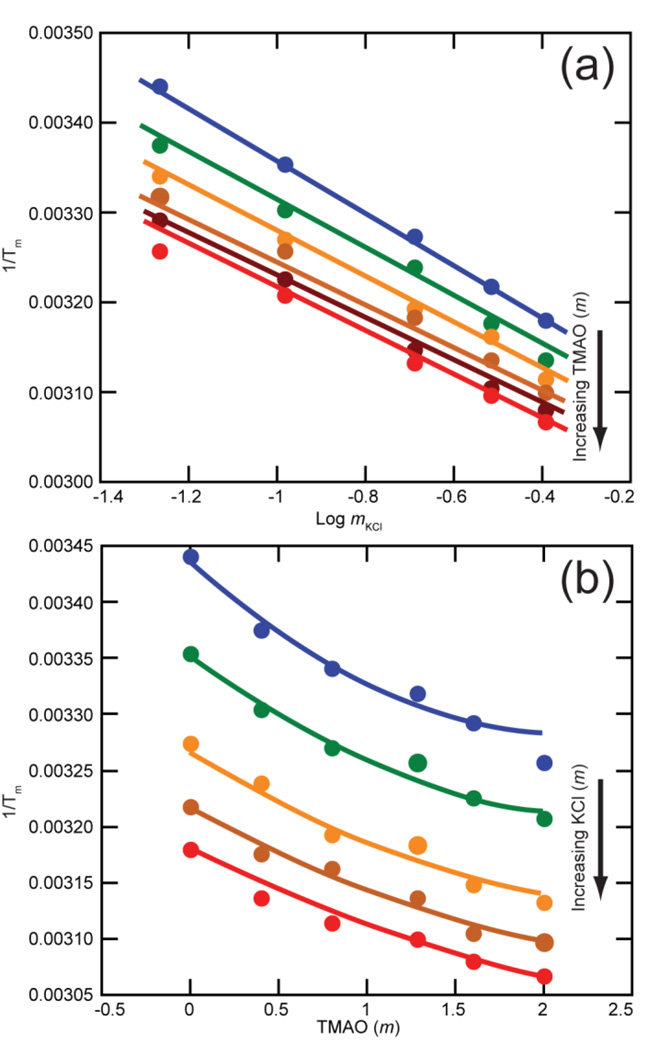
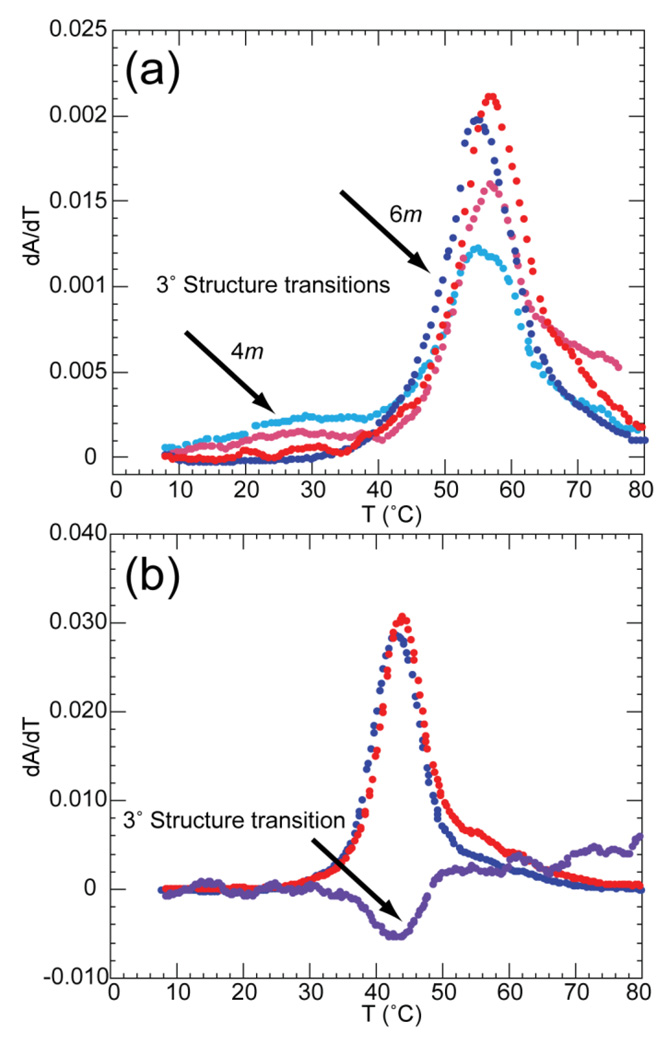
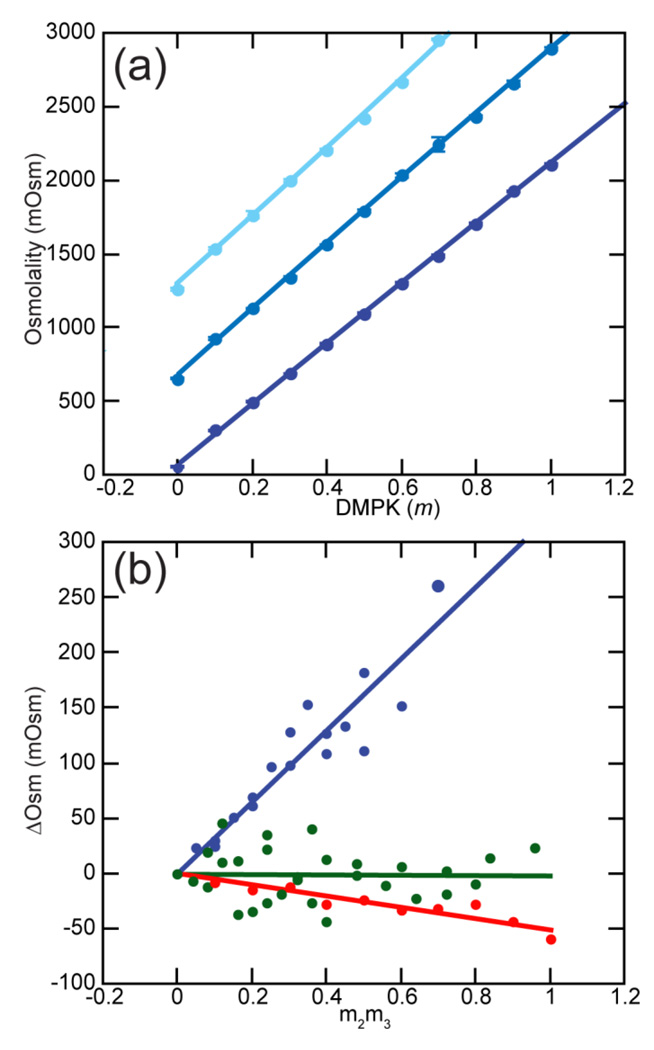
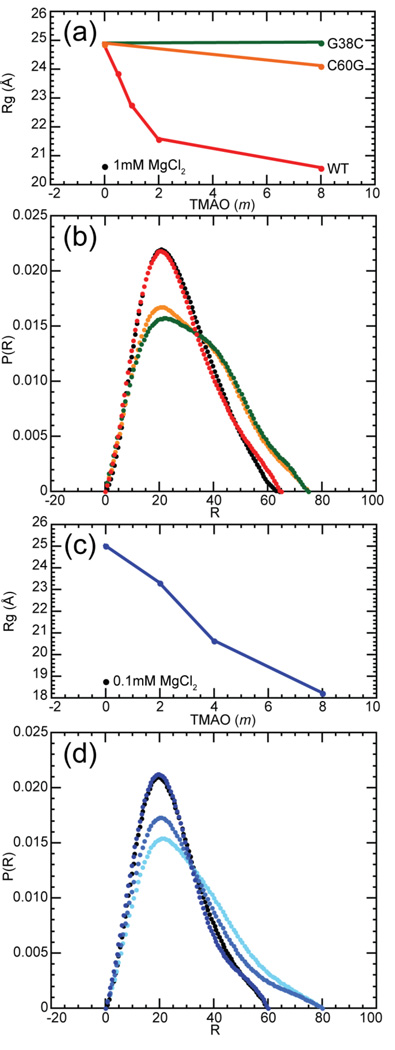
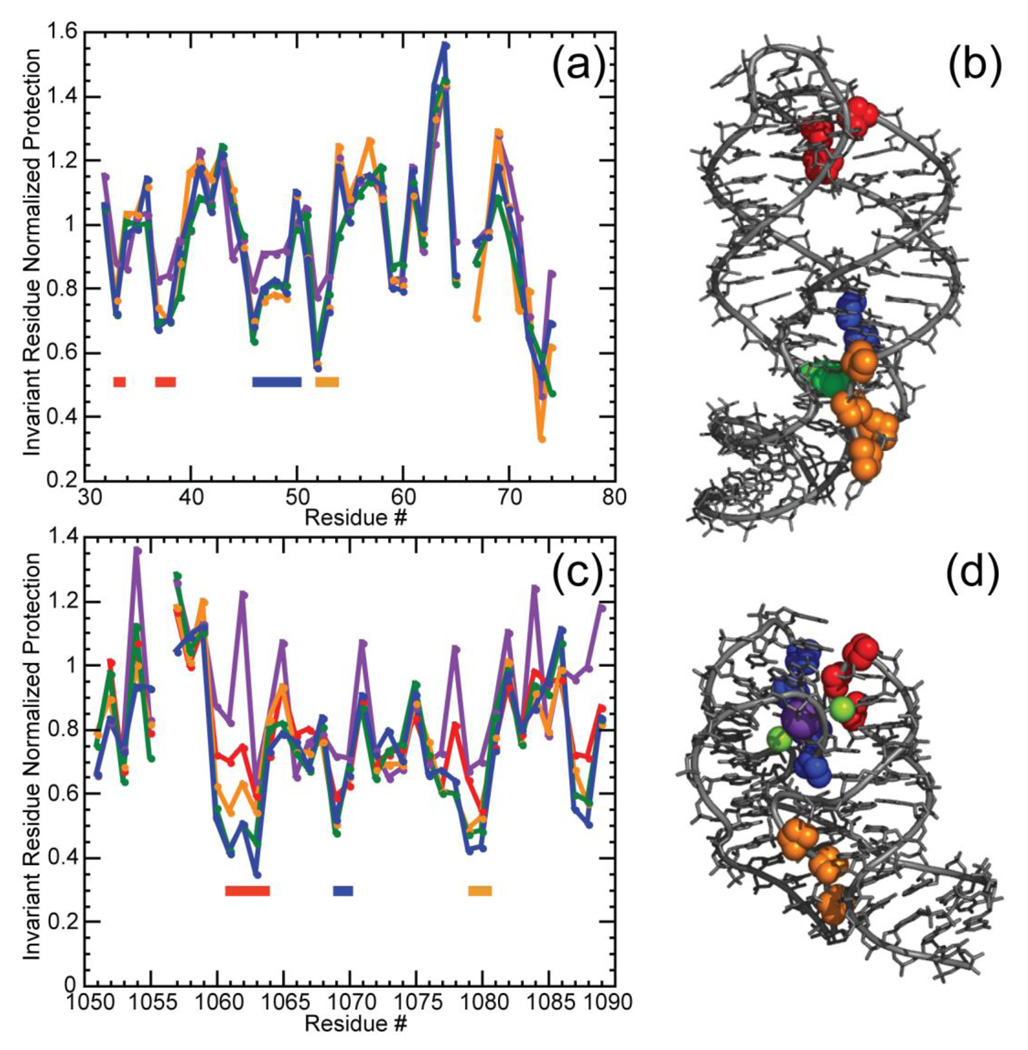
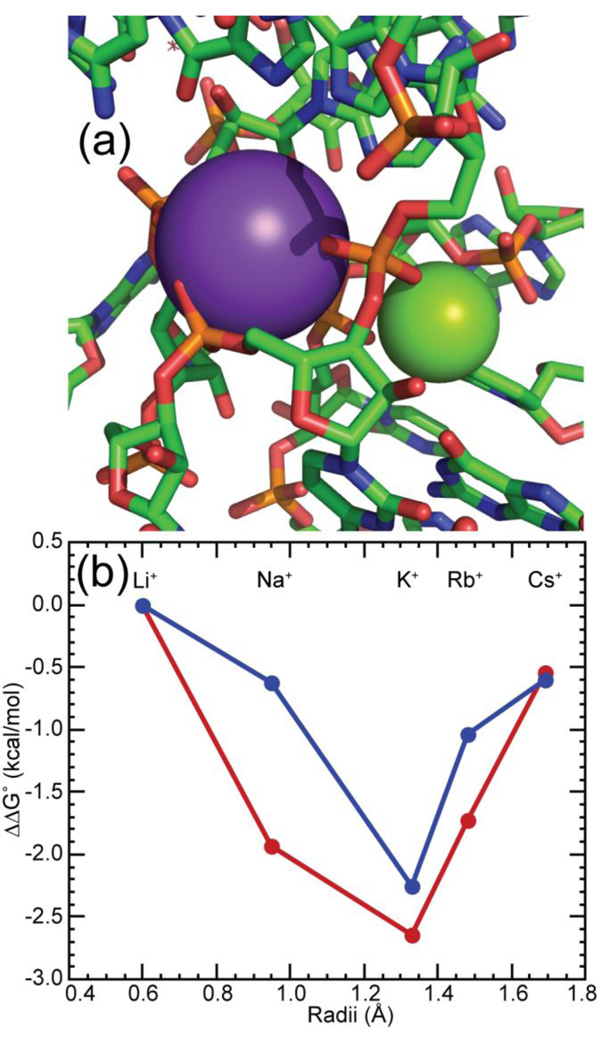
The stabilization of RNA tertiary structures by ions is well known, but the neutral osmolyte trimethylamine oxide (TMAO) can also effectively stabilize RNA tertiary structure. To begin to understand the physical basis for the effects of TMAO on RNA, we have quantitated the TMAO-induced stabilization of five RNAs with known structures. So-called m values, the increment in unfolding free energy per molal of osmolyte at constant KCl activity, are ∼0 for a hairpin secondary structure and between 0.70 and 1.85 kcal mol(-1)m(-1) for four RNA tertiary structures (30-86 nt). Further analysis of two RNAs by small-angle X-ray scattering and hydroxyl radical probing shows that TMAO reduces the radius of gyration of the unfolded ensemble to the same endpoint as seen in titration with Mg(2+) and that the structures stabilized by TMAO and Mg(2+) are indistinguishable. Remarkably, TMAO induces the native conformation of a Mg(2+) ion chelation site formed in part by a buried phosphate, even though Mg(2+) is absent. TMAO interacts weakly, if at all, with KCl, ruling out the possibility that TMAO stabilizes RNA indirectly by increasing salt activity. TMAO is, however, strongly excluded from the vicinity of dimethylphosphate (unfavorable interaction free energy, +211 cal mol(-1)m(-1) for the potassium salt), an ion that mimics the RNA backbone phosphate. We suggest that formation of RNA tertiary structure is accompanied by substantial phosphate dehydration (loss of 66-173 water molecules in the RNA structures studied) and that TMAO works principally by reducing the energetic penalty associated with this dehydration. The strong parallels we find between the effects of TMAO and Mg(2+) suggest that RNA sequence is more important than specific ion interactions in specifying the native structure.
RNA 三级结构的稳定是众所周知的,但是中性渗透调节剂三甲胺氧化物(TMAO)也可以有效地稳定 RNA 三级结构。为了开始理解 TMAO 对 RNA 的影响的物理基础,我们已经定量了五种具有已知结构的 RNA 的 TMAO 诱导稳定性。所谓的 m 值,即在恒定 KCl 活度下每摩尔渗透压摩尔的渗透调节剂引起的解折叠自由能增量,对于发夹二级结构约为 0,对于四个 RNA 三级结构(30-86nt)则在 0.70 到 1.85 kcal mol(-1)m(-1)之间。通过小角 X 射线散射和羟基自由基探测对两种 RNA 的进一步分析表明,TMAO 将展开的整体的回转半径减小到与用 Mg(2+)滴定相同的终点,并且 TMAO 和 Mg(2+)稳定的结构无法区分。值得注意的是,TMAO 诱导了由部分埋藏的磷酸盐形成的 Mg(2+)离子螯合位点的天然构象,即使没有 Mg(2+)。TMAO 与 KCl 的相互作用很弱,如果有的话,排除了 TMAO 通过增加盐活性间接稳定 RNA 的可能性。然而,TMAO 强烈地被排除在二甲基磷酸盐(不利的相互作用自由能,对于钾盐为+211 cal mol(-1)m(-1))的附近,二甲基磷酸盐模拟 RNA 骨架磷酸盐。我们认为 RNA 三级结构的形成伴随着大量磷酸盐的去水合作用(在所研究的 RNA 结构中损失 66-173 个水分子),并且 TMAO 的主要作用是降低与这种去水合作用相关的能量惩罚。我们发现 TMAO 和 Mg(2+)的作用之间存在很强的相似性,这表明 RNA 序列比特定离子相互作用更重要,以指定天然结构。