Division of Basic Medical Sciences, St. George's, University of London, Cranmer Terrace, London SW17 0RE, UK.
J Biol Chem. 2011 Apr 8;286(14):12308-16. doi: 10.1074/jbc.M110.157057. Epub 2011 Feb 7.
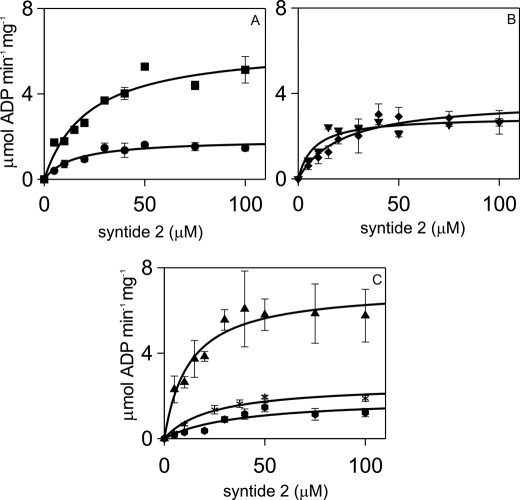
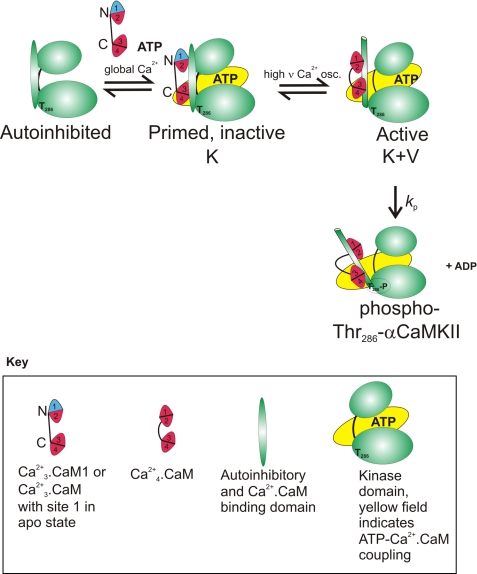
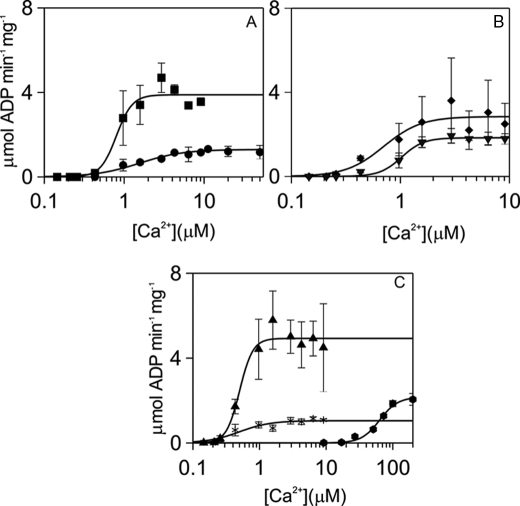
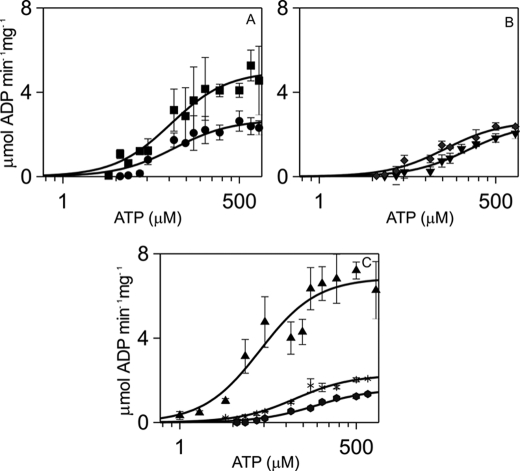
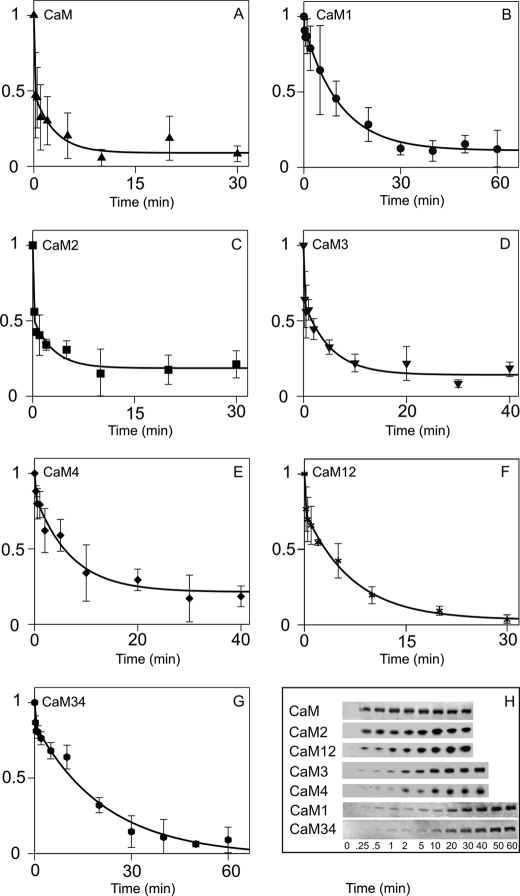
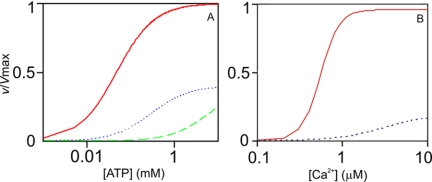
N-methyl-D-aspartic acid receptor-dependent long term potentiation (LTP), a model of memory formation, requires Ca2+·calmodulin-dependent protein kinase II (αCaMKII) activity and Thr286 autophosphorylation via both global and local Ca2+ signaling, but the mechanisms of signal transduction are not understood. We tested the hypothesis that the Ca2+-binding activator protein calmodulin (CaM) is the primary decoder of Ca2+ signals, thereby determining the output, e.g. LTP. Thus, we investigated the function of CaM mutants, deficient in Ca2+ binding at sites 1 and 2 of the N-terminal lobe or sites 3 and 4 of the C-terminal CaM lobe, in the activation of αCaMKII. Occupancy of CaM Ca2+ binding sites 1, 3, and 4 is necessary and sufficient for full activation. Moreover, the N- and C-terminal CaM lobes have distinct functions. Ca2+ binding to N lobe Ca2+ binding site 1 increases the turnover rate of the enzyme 5-fold, whereas the C lobe plays a dual role; it is required for full activity, but in addition, via Ca2+ binding site 3, it stabilizes ATP binding to αCaMKII 4-fold. Thr286 autophosphorylation is also dependent on Ca2+ binding sites on both the N and the C lobes of CaM. As the CaM C lobe sites are populated by low amplitude/low frequency (global) Ca2+ signals, but occupancy of N lobe site 1 and thus activation of αCaMKII requires high amplitude/high frequency (local) Ca2+ signals, lobe-specific sensing of Ca2+-signaling patterns by CaM is proposed to explain the requirement for both global and local Ca2+ signaling in the induction of LTP via αCaMKII.
N-甲基-D-天冬氨酸受体依赖性长时程增强(LTP)是记忆形成的模型,需要 Ca2+·钙调蛋白依赖性蛋白激酶 II(αCaMKII)活性和 Thr286 通过全局和局部 Ca2+信号的自身磷酸化,但信号转导的机制尚不清楚。我们检验了这样一个假设,即 Ca2+结合激活蛋白钙调蛋白(CaM)是 Ca2+信号的主要解码器,从而决定了输出,例如 LTP。因此,我们研究了 CaM 突变体的功能,这些突变体在 N 端结构域的 1 和 2 位或 C 端 CaM 结构域的 3 和 4 位的 Ca2+结合位点缺乏 Ca2+结合,在 αCaMKII 的激活中的作用。CaM Ca2+结合位点 1、3 和 4 的占据对于完全激活是必需和充分的。此外,N 和 C 端 CaM 结构域具有不同的功能。Ca2+结合到 N 结构域的 Ca2+结合位点 1 会使酶的周转率增加 5 倍,而 C 结构域则具有双重作用;它是完全活性所必需的,但此外,通过 Ca2+结合位点 3,它将 ATP 与 αCaMKII 的结合稳定 4 倍。Thr286 自身磷酸化也依赖于 CaM 的 N 和 C 结构域上的 Ca2+结合位点。由于 CaM C 结构域的位点被低幅度/低频(全局)Ca2+信号占据,而 N 结构域位点 1 的占据和因此 αCaMKII 的激活需要高幅度/高频(局部)Ca2+信号,因此建议 CaM 对 Ca2+-信号模式的叶特异性感知可以解释在通过 αCaMKII 诱导 LTP 中全局和局部 Ca2+信号的双重需求。