Affentranger Sarah, Martinelli Sibylla, Hahn Jonas, Rossy Jérémie, Niggli Verena
Dept. of Pathology, University of Bern, CH-3010 Bern, Switzerland.
BMC Cell Biol. 2011 Jun 22;12:28. doi: 10.1186/1471-2121-12-28.
Different types of membrane microdomains (rafts) have been postulated to be present in the rear and front of polarized migrating T-lymphocytes. Disruption of rafts by cholesterol sequestration prevents T-cell polarization and migration. Reggie/flotillin-1 and -2 are two highly homologous proteins that are thought to shape membrane microdomains. We have previously demonstrated the enrichment of flotillins in the uropod of human neutrophils. We have now investigated mechanisms involved in chemokine-induced flotillin reorganization in human T-lymphocytes, and possible roles of flotillins in lymphocyte polarization.
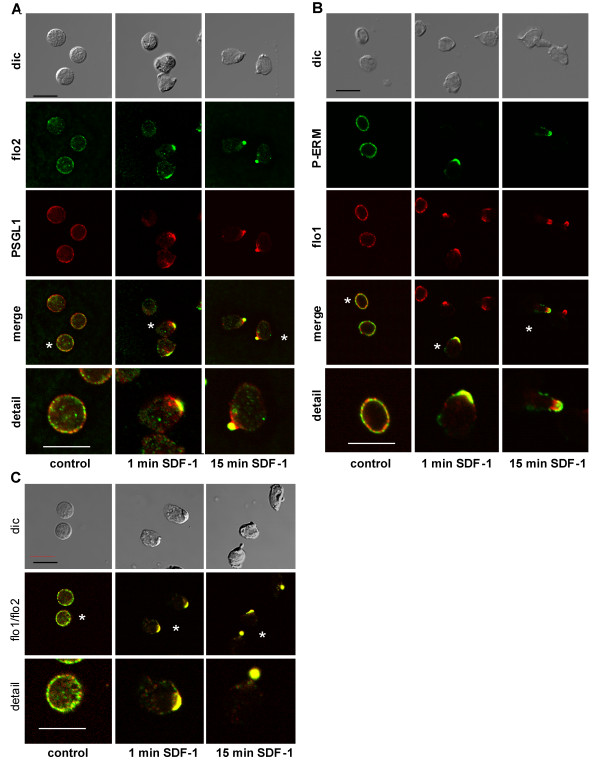
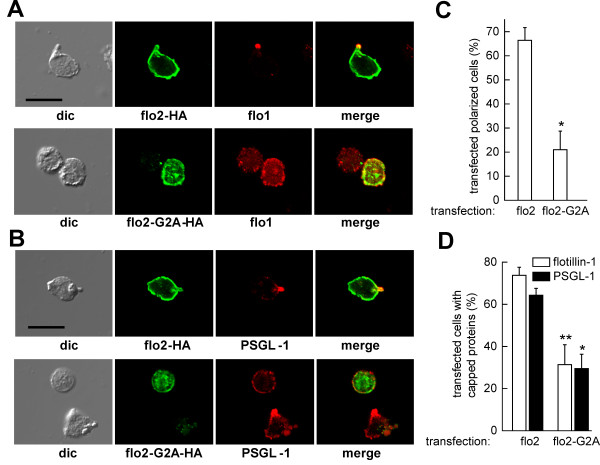
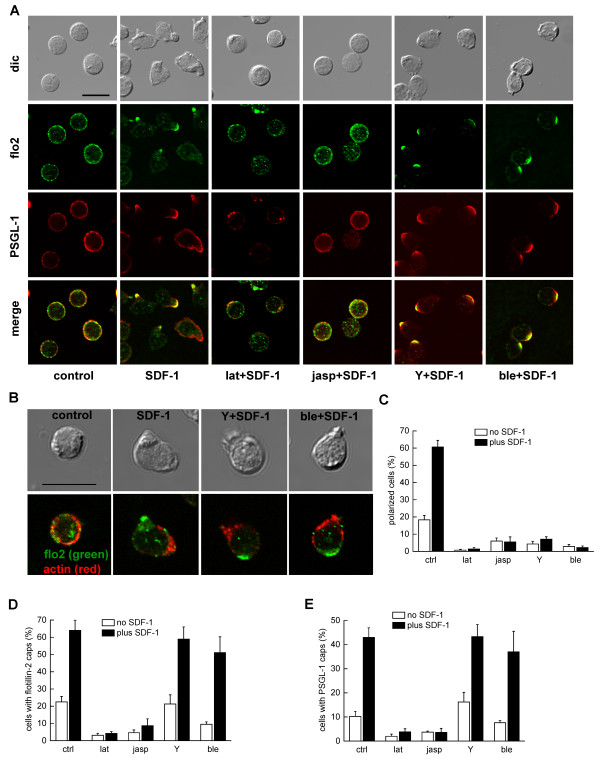
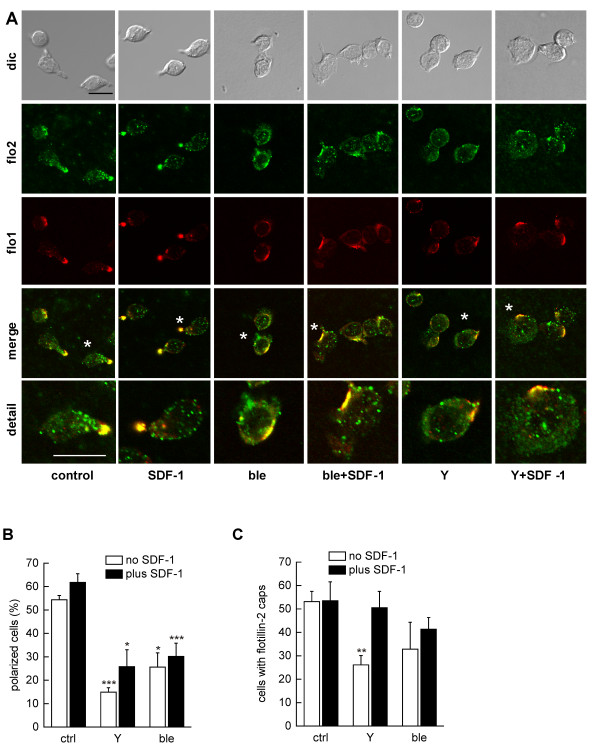
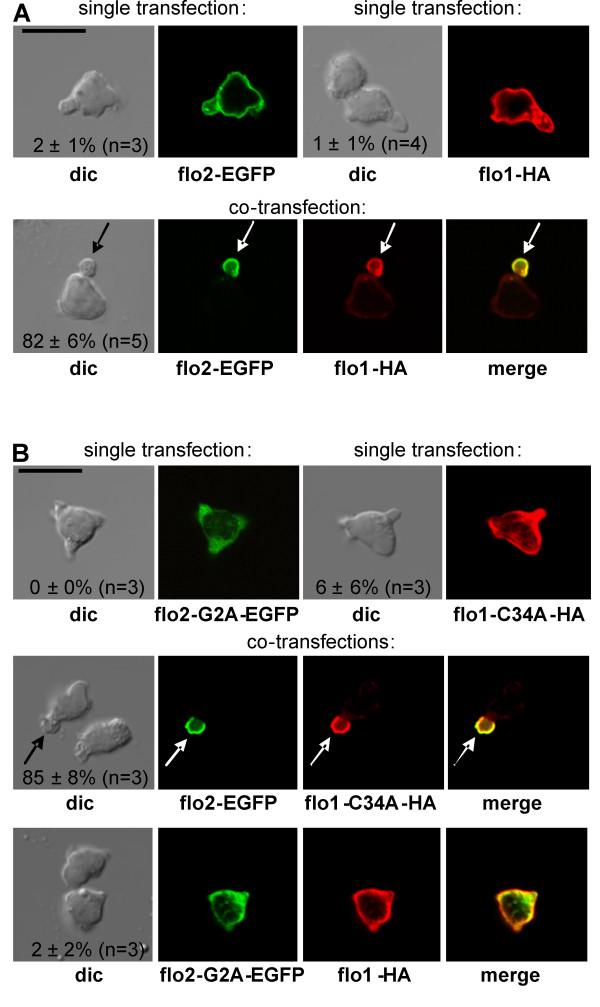
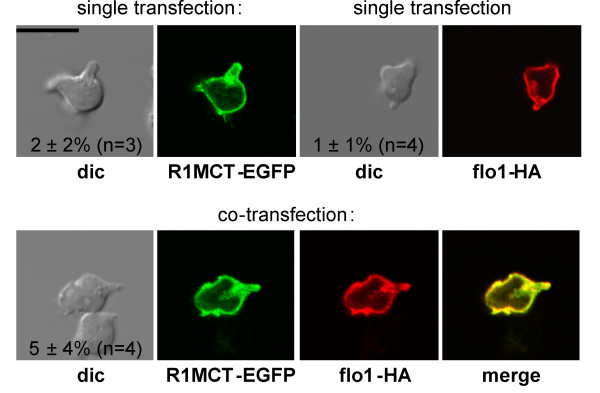
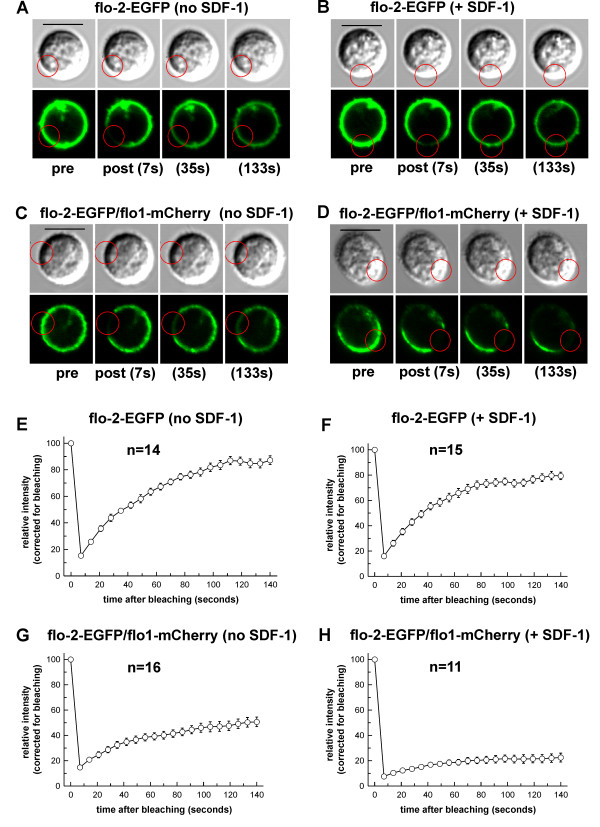
We studied flotillin reorganization and lateral mobility at the plasma membrane using immunofluorescence staining and FRAP (fluorescence recovery after photobleaching). We show that flotillins redistribute early upon chemokine stimulation, and form very stable caps in the uropods of human peripheral blood T-lymphocytes, colocalizing with the adhesion molecule PSGL-1 and activated ezrin/radixin/moesin (ERM) proteins. Chemokine-induced formation of stable flotillin caps requires integrity and dynamics of the actin cytoskeleton, but is not abolished by inhibitors suppressing Rho-kinase or myosin II activity. Tagged flotillin-2 and flotillin-1 coexpressed in T-lymphocytes, but not singly expressed proteins, colocalize in stable caps at the tips of uropods. Lateral mobility of coexpressed flotillins at the plasma membrane is already partially restricted in the absence of chemokine. Incubation with chemokine results in almost complete immobilization of flotillins. Capping is abolished when wild-type flotillin-1 is coexpressed with a mutant of flotillin-2 (G2A) that is unable to interact with the plasma membrane, or with a deletion mutant of flotillin-2 that lacks a putative actin-binding domain. Wild-type flotillin-2 in contrast forms caps when coexpressed with a mutant of flotillin-1 unable to interact with membranes. Transfection of T-lymphocytes with flotillin-2-G2A reduces cell polarization and uropod recruitment of endogenous flotillin-1 and PSGL-1.
Our data suggest that stable flotillin cap formation in the rear of polarized T-lymphocytes requires flotillin heterooligomer formation, as well as direct F-actin interactions of flotillin-2 and raft/membrane association of flotillin-2, but not -1. Our data also implicate flotillin-rich actin-dependent membrane microdomains in T-lymphocyte uropod formation.
据推测,不同类型的膜微结构域(脂筏)存在于极化迁移的T淋巴细胞的后部和前部。通过胆固醇螯合破坏脂筏会阻止T细胞极化和迁移。Reggie/小窝蛋白-1和-2是两种高度同源的蛋白质,被认为能够塑造膜微结构域。我们之前已经证明小窝蛋白在人中性粒细胞的尾足中富集。我们现在研究了趋化因子诱导人T淋巴细胞中小窝蛋白重组的机制,以及小窝蛋白在淋巴细胞极化中的可能作用。
我们使用免疫荧光染色和光漂白后荧光恢复(FRAP)技术研究了质膜上的小窝蛋白重组和侧向流动性。我们发现,趋化因子刺激后,小窝蛋白会早期重新分布,并在人外周血T淋巴细胞的尾足中形成非常稳定的帽状物,与黏附分子PSGL-1和活化的埃兹蛋白/根蛋白/膜突蛋白(ERM)共定位。趋化因子诱导的稳定小窝蛋白帽状物的形成需要肌动蛋白细胞骨架的完整性和动态性,但抑制Rho激酶或肌球蛋白II活性的抑制剂并不能消除这种现象。在T淋巴细胞中共表达的带标签的小窝蛋白-2和小窝蛋白-1会共定位在尾足尖端的稳定帽状物中,而单独表达的蛋白质则不会。在没有趋化因子的情况下,共表达的小窝蛋白在质膜上的侧向流动性已经部分受限。用趋化因子孵育会导致小窝蛋白几乎完全固定。当野生型小窝蛋白-1与无法与质膜相互作用的小窝蛋白-2突变体(G2A)或缺乏假定肌动蛋白结合结构域的小窝蛋白-2缺失突变体共表达时,帽状物形成被消除。相反,当野生型小窝蛋白-2与无法与膜相互作用的小窝蛋白-1突变体共表达时,会形成帽状物。用小窝蛋白-2-G2A转染T淋巴细胞会降低细胞极化以及内源性小窝蛋白-1和PSGL-1的尾足募集。
我们的数据表明,极化T淋巴细胞后部稳定的小窝蛋白帽状物形成需要小窝蛋白异源寡聚体的形成,以及小窝蛋白-2与F-肌动蛋白的直接相互作用和小窝蛋白-2与脂筏/膜的结合,但小窝蛋白-1不需要。我们的数据还表明富含小窝蛋白的肌动蛋白依赖性膜微结构域参与了T淋巴细胞尾足的形成。