Wayengera Misaki
Unit of Genetics, Genomics & Theoretical Biology, Dept of Pathology, School of Biomedical Science, College of Health Sciences, Makerere University, P O Box 7072 Kampala, Uganda.
Theor Biol Med Model. 2011 Jun 24;8:23. doi: 10.1186/1742-4682-8-23.
Herpes simplex type II (HSV-2) is a member of the family herpesviridae. Human infection with this double stranded linear DNA virus causes genital ulcerative disease and existing treatment options only serve to resolve the symptomatology (ulcers) associated with active HSV-2 infection but do not eliminate latent virus. As a result, infection with HSV-2 follows a life-long relapsing (active versus latent) course. On the basis of a primitive bacterium anti-phage DNA defense, the restriction modification (R-M) system, we previously identified the Escherichia coli restriction enzyme (REase) EcoRII as a novel peptide to excise or irreversibly disrupt latent HSV-2 DNA from infected cells. However, sequences of the site specificity palindrome of EcoRII 5'-CCWGG-3' (W = A or T) are equally present within the human genome and are a potential source of host-genome toxicity. This feature has limited previous HSV-2 EcoRII based therapeutic models to microbicides only, and highlights the need to engineer artificial REases (zinc finger nucleases-ZFNs) with specificity to HSV-2 genomic-DNA only. Herein, the therapeutic-potential of zinc finger arrays (ZFAs) and ZFNs is identified and modeled, with unique specificity to the HSV-2 genome.
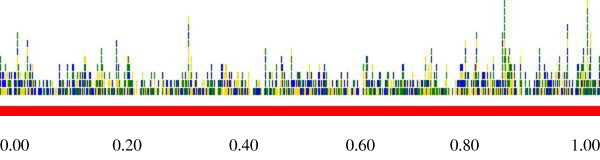
Using the whole genome of HSV-2 strain HG52 (Dolan A et al.,), and with the ZFN-consortium's CoDA-ZiFiT software pre-set at default, more than 28,000 ZFAs with specificity to HSV-2 DNA were identified. Using computational assembly (through in-silico linkage to the Flavobacterium okeanokoites endonuclease Fok I of the type IIS class), 684 ZFNs with specificity to the HSV-2 genome, were constructed. Graphic-analysis of the HSV-2 genome-cleavage pattern using the afore-identified ZFNs revealed that the highest cleavage-incidence occurred within the 30,950 base-pairs (~between the genomic context coordinates 0.80 and 1.00) at the 3' end of the HSV-2 genome. At approximately 3,095 bp before and after the 5' and 3' ends of the HSV-2 genome (genomic context coordinates 0.02 and 0.98, respectively) were specificity sites of ZFNs suited for the complete excision of over 60% of HSV-2 genomic material from within infected human cells, through the process of non-homologous end joining (NHEJ). Furthermore, a model concerning a recombinant (ICP10-PK mutant) replication competent HSV-2 viral vector for delivering and transducing a diploid copy (or pair) of the HSV-2-genome-specific ZFN genotype within neuronal tissue, is presented.
ZFNs with specificity to HSV-2 genomic DNA that are precursors of novel host-genome expressed HSV-2 gene-therapeutics or vaccines were identified.
单纯疱疹病毒II型(HSV - 2)是疱疹病毒科的成员。人类感染这种双链线性DNA病毒会导致生殖器溃疡性疾病,现有的治疗方法仅用于缓解与活动性HSV - 2感染相关的症状(溃疡),但不能消除潜伏病毒。因此,HSV - 2感染呈终身复发(活动期与潜伏期交替)病程。基于一种原始细菌的抗噬菌体DNA防御机制——限制修饰(R - M)系统,我们之前鉴定出大肠杆菌限制酶(REase)EcoRII是一种新型肽,可从感染细胞中切除或不可逆地破坏潜伏的HSV - 2 DNA。然而,EcoRII的位点特异性回文序列5'-CCWGG-3'(W = A或T)在人类基因组中也同样存在,是宿主基因组毒性的潜在来源。这一特性使得先前基于HSV - 2 EcoRII的治疗模型仅局限于杀微生物剂,并凸显了设计仅对HSV - 2基因组DNA具有特异性的人工限制酶(锌指核酸酶 - ZFNs)的必要性。在此,我们鉴定并模拟了锌指阵列(ZFAs)和ZFNs对HSV - 2基因组具有独特特异性的治疗潜力。
使用HSV - 2毒株HG52的全基因组(Dolan A等人),并将ZFN联盟的CoDA - ZiFiT软件预设为默认值,鉴定出了28,000多个对HSV - 2 DNA具有特异性的ZFA。通过计算组装(通过计算机模拟与IIS类的海洋黄杆菌内切酶Fok I连接),构建了684个对HSV - 2基因组具有特异性的ZFN。使用上述鉴定出的ZFN对HSV - 2基因组切割模式进行图形分析,结果显示最高切割发生率出现在HSV - 2基因组3'端的30,950个碱基对处(约在基因组上下文坐标0.80和1.00之间)。在HSV - 2基因组5'端和3'端之前和之后大约3,095 bp处(分别为基因组上下文坐标0.02和0.98)是ZFN的特异性位点,通过非同源末端连接(NHEJ)过程,这些位点适合从感染的人类细胞中完全切除超过60%的HSV - 2基因组物质。此外,还提出了一个关于重组(ICP10 - PK突变体)复制缺陷型HSV - 2病毒载体的模型,该载体用于在神经元组织中递送和转导HSV - 2基因组特异性ZFN基因型的二倍体拷贝(或一对)。
鉴定出了对HSV - 2基因组DNA具有特异性的ZFN,它们是新型宿主基因组表达的HSV - 2基因治疗或疫苗的前体。